

Interacting targets of the farnesyl of transducin gamma-subunit
- PMID: 18636747
- PMCID: PMC2646881
- DOI: 10.1021/bi800359h
Interacting targets of the farnesyl of transducin gamma-subunit
Abstract
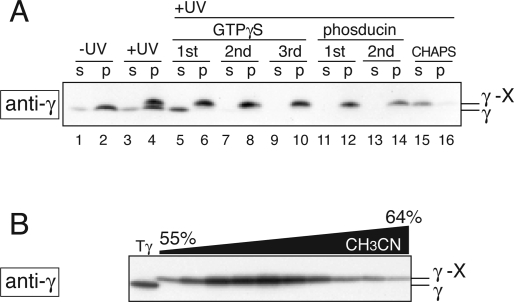
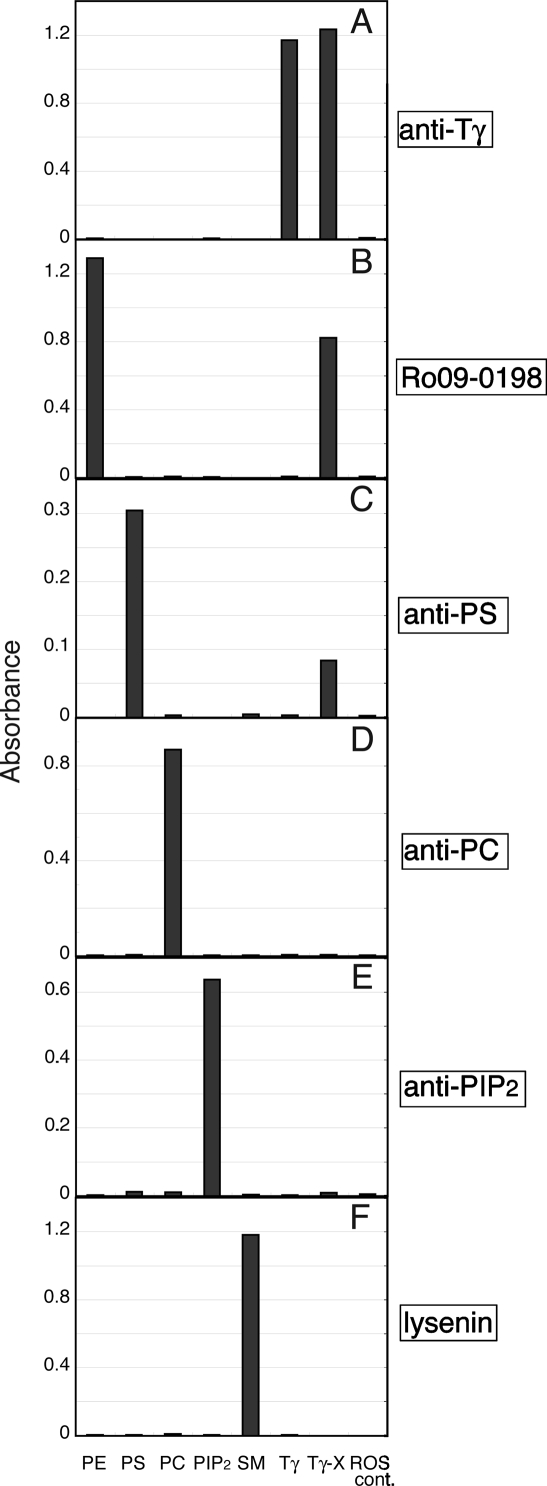
G protein gamma-subunits are isoprenylated and carboxyl methylated at the C-terminal cysteine residue, and the set of the posttranslational modifications is indispensable for the function of the photoreceptor G protein transducin (Talpha/Tbetagamma). To explore farnesyl-mediated molecular interactions, we investigated molecular targets of a Tbetagamma analogue that was engineered to have a photoreactive farnesyl analogue, (3-azidophenoxy)geranyl (POG), covalently bound to the C-terminal cysteine of Tgamma. POG-modified Tbetagamma was further subjected to modification by methylation at the C-terminal carboxyl group, which copies a complete set of the known posttranscriptional modifications of Tbetagamma. Photoaffinity labeling experiment with the photoreactive Tbetagamma analogue in its free form indicated that the POG moiety of Tgamma interacted with Tbeta. In the trimeric Talpha/Tbetagamma complex, the POG moiety was cross-linked with Talpha in addition to concurrent affinity labeling of Tbeta. When photoreactive Tbetagamma was reconstituted with Talpha and light-activated rhodopsin (Rh*) in rod outer segment (ROS) membranes, the POG moiety interacted with not only Talpha and Tbeta but also Rh* and membrane phospholipids. The cross-linked phospholipid species was analyzed by ELISA employing a variety of lipid-binding probes, which revealed phosphatidylethanolamine (PE) and phosphatidylserine (PS) as selective phospholipids for POG interaction in the ROS membranes. These results demonstrate that the modifying group of Tgamma plays an active role in protein-protein and protein-membrane interactions and suggest that the farnesyl-PE/PS interaction may support the efficient formation of the signaling ternary complex between transducin and Rh*.
Figures
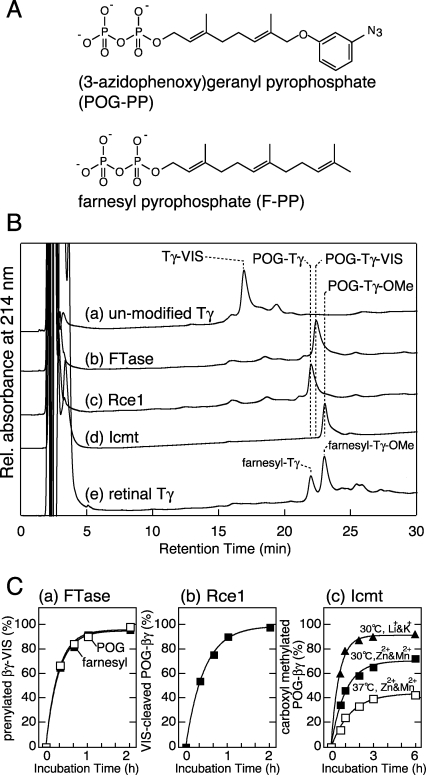
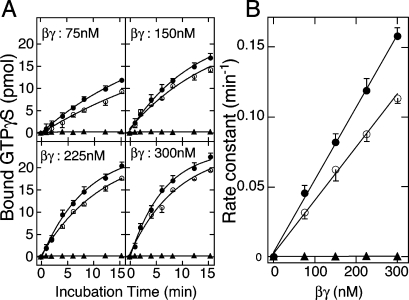
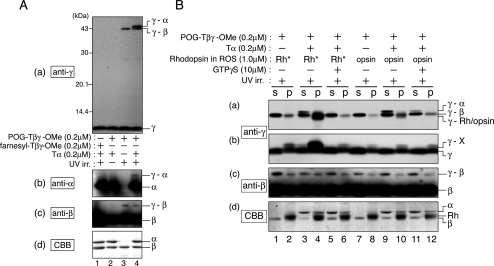
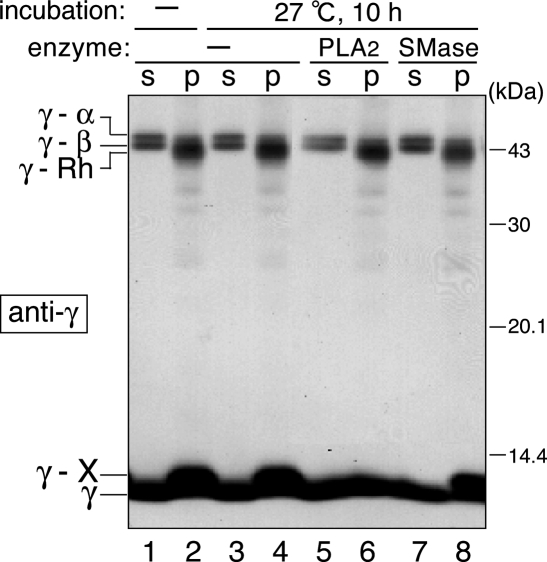
References
-
- Arshavsky V. Y.; Lamb T. D.; Pugh E. N. Jr. (2002) G proteins and phototransduction. Annu. Rev. Physiol. 64, 153–187. - PubMed
-
- Fukada Y.; Takao T.; Ohguro H.; Yoshizawa T.; Akino T.; Shimonishi Y. (1990) Farnesylated γ-subunit of photoreceptor G protein indispensable for GTP-binding. Nature 346, 658–660. - PubMed
-
- Matsuda T.; Fukada Y. (2000) Functional analysis of farnesylation and methylation of transducin. Methods Enzymol. 316, 465–481. - PubMed
-
- Casey P. J.; Seabra M. C. (1996) Protein prenyltransferases. J. Biol. Chem. 271, 5289–5292. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
