

Identification of residues in the Cmu4 domain of polymeric IgM essential for interaction with Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1)
- PMID: 18641336
- PMCID: PMC2696179
- DOI: 10.4049/jimmunol.181.3.1988
Identification of residues in the Cmu4 domain of polymeric IgM essential for interaction with Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1)
Abstract
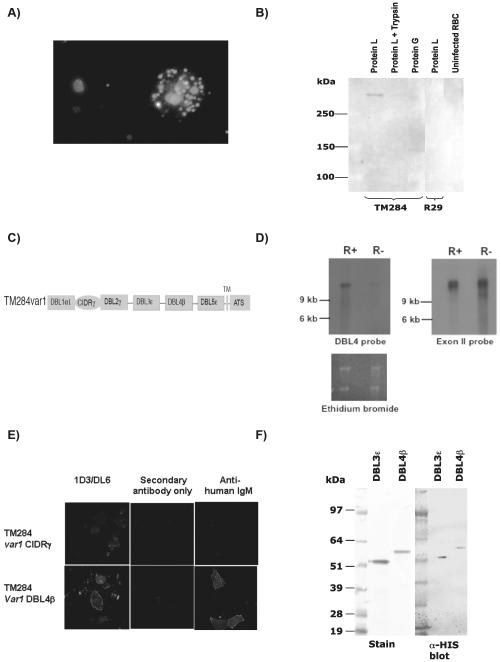
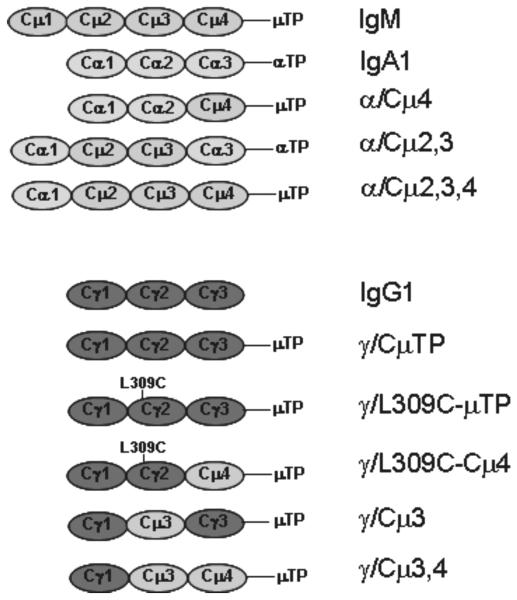
The binding of nonspecific human IgM to the surface of infected erythrocytes is important in rosetting, a major virulence factor in the pathogenesis of severe malaria due to Plasmodium falciparum, and IgM binding has also been implicated in placental malaria. Herein we have identified the IgM-binding parasite ligand from a virulent P. falciparum strain as PfEMP1 (TM284var1 variant), and localized the region within this PfEMP1 variant that binds IgM (DBL4beta domain). We have used this parasite IgM-binding protein to investigate the interaction with human IgM. Interaction studies with domain-swapped Abs, IgM mutants, and anti-IgM mAbs showed that PfEMP1 binds to the Fc portion of the human IgM H chain and requires the IgM Cmu4 domain. Polymerization of IgM was shown to be crucial for the interaction because PfEMP1 binding did not occur with mutant monomeric IgM molecules. These results with PfEMP1 protein have physiological relevance because infected erythrocytes from strain TM284 and four other IgM-binding P. falciparum strains showed analogous results to those seen with the DBL4beta domain. Detailed investigation of the PfEMP1 binding site on IgM showed that some of the critical amino acids in the IgM Cmu4 domain are equivalent to those regions of IgG and IgA recognized by Fc-binding proteins from bacteria, suggesting that this region of Ig molecules may be of major functional significance in host-microbe interactions. We have therefore shown that PfEMP1 is an Fc-binding protein of malaria parasites specific for polymeric human IgM, and that it shows functional similarities with Fc-binding proteins from pathogenic bacteria.
Figures






Similar articles
-
Investigating the function of Fc-specific binding of IgM to Plasmodium falciparum erythrocyte membrane protein 1 mediating erythrocyte rosetting.Cell Microbiol. 2015 Jun;17(6):819-31. doi: 10.1111/cmi.12403. Epub 2015 Jan 28. Cell Microbiol. 2015. PMID: 25482886 Free PMC article.
-
Identification of the minimal binding region of a Plasmodium falciparum IgM binding PfEMP1 domain.Mol Biochem Parasitol. 2015 May;201(1):76-82. doi: 10.1016/j.molbiopara.2015.06.001. Epub 2015 Jun 18. Mol Biochem Parasitol. 2015. PMID: 26094597 Free PMC article.
-
Multiple Plasmodium falciparum Erythrocyte Membrane Protein 1 Variants per Genome Can Bind IgM via Its Fc Fragment Fcμ.Infect Immun. 2015 Oct;83(10):3972-81. doi: 10.1128/IAI.00337-15. Epub 2015 Jul 27. Infect Immun. 2015. PMID: 26216422 Free PMC article.
-
Decoding the language of var genes and Plasmodium falciparum sequestration.Trends Parasitol. 2001 Nov;17(11):538-45. doi: 10.1016/s1471-4922(01)02079-7. Trends Parasitol. 2001. PMID: 11872399 Review.
-
PfEMP1 - A Parasite Protein Family of Key Importance in Plasmodium falciparum Malaria Immunity and Pathogenesis.Adv Parasitol. 2015 Apr;88:51-84. doi: 10.1016/bs.apar.2015.02.004. Epub 2015 Mar 23. Adv Parasitol. 2015. PMID: 25911365 Review.
Cited by
-
IgM, Fc mu Rs, and malarial immune evasion.J Immunol. 2010 May 1;184(9):4597-603. doi: 10.4049/jimmunol.1000203. J Immunol. 2010. PMID: 20410497 Free PMC article. Review.
-
Variant surface antigens of Plasmodium falciparum and their roles in severe malaria.Nat Rev Microbiol. 2017 Aug;15(8):479-491. doi: 10.1038/nrmicro.2017.47. Epub 2017 Jun 12. Nat Rev Microbiol. 2017. PMID: 28603279 Review.
-
The role of PfEMP1 as targets of naturally acquired immunity to childhood malaria: prospects for a vaccine.Parasitology. 2016 Feb;143(2):171-86. doi: 10.1017/S0031182015001274. Epub 2016 Jan 7. Parasitology. 2016. PMID: 26741401 Free PMC article. Review.
-
Adhesion of Plasmodium falciparum-infected erythrocytes to human cells: molecular mechanisms and therapeutic implications.Expert Rev Mol Med. 2009 May 26;11:e16. doi: 10.1017/S1462399409001082. Expert Rev Mol Med. 2009. PMID: 19467172 Free PMC article. Review.
-
Detection of naturally acquired, strain-transcending antibodies against rosetting Plasmodium falciparum strains in humans.Infect Immun. 2024 Jul 11;92(7):e0001524. doi: 10.1128/iai.00015-24. Epub 2024 Jun 6. Infect Immun. 2024. PMID: 38842304 Free PMC article.
References
-
- Burton DR. Structure and function of antibodies. In: Calabi F, Neuberger MS, editors. Molecular Genetics of Immunoglobulin. Elsevier Science Publishers B.V. (Biomedical Division); 1987. pp. 1–49.
-
- Perkins SJ, Nealis AS, Sutton BJ, J B, Feinstein A. Solution structure of human and mouse immunoglobulin M by synchrotron X-ray scattering and molecular graphics modelling. A possible mechanism for complement activation. J. Mol. Biol. 1991;221:1345–1366. - PubMed
-
- Shibuya A, Sakamoto N, Shimizu Y, Shibuya K, Osawa M, Hiroyama T, Eyre HJ, Sutherland GR, Endo Y, Fujita T, et al. Fc alpha/mu receptor mediates endocytosis of IgM-coated microbes. Nat. Immunol. 2000;1:441–446. - PubMed
-
- Sakamoto N, Shibuya K, Shimizu Y, Yotsumoto K, Miyabayashi T, Sakano S, Tsuji T, Nakayama E, Nakauchi H, Shibuya A. A novel Fc receptor for IgA and IgM is expressed on both hematopoietic and non-hematopoietic tissues. Eur. J. Immunol. 2001;31:1310–1316. - PubMed
-
- Cho Y, Usui K, Honda S, Tahara-Hanaoka S, Shibuya K, Shibuya A. Molecular characteristics of IgA and IgM Fc binding to the Fcalpha/muR. Biochem. Biophys. Res. Commun. 2006;345:474–478. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
