

Opposing roles of p190RhoGAP and Ect2 RhoGEF in regulating cytokinesis
- PMID: 18642445
- PMCID: PMC2791401
- DOI: 10.4161/cc.7.13.6128
Opposing roles of p190RhoGAP and Ect2 RhoGEF in regulating cytokinesis
Abstract
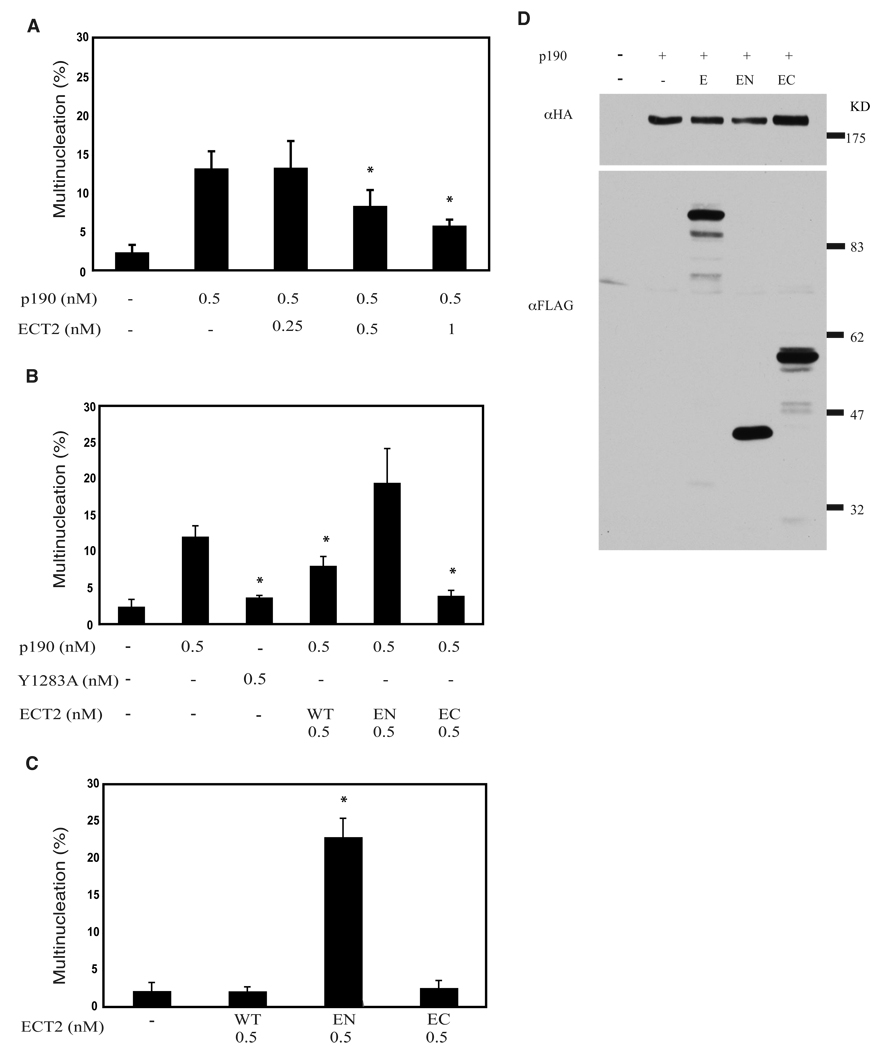
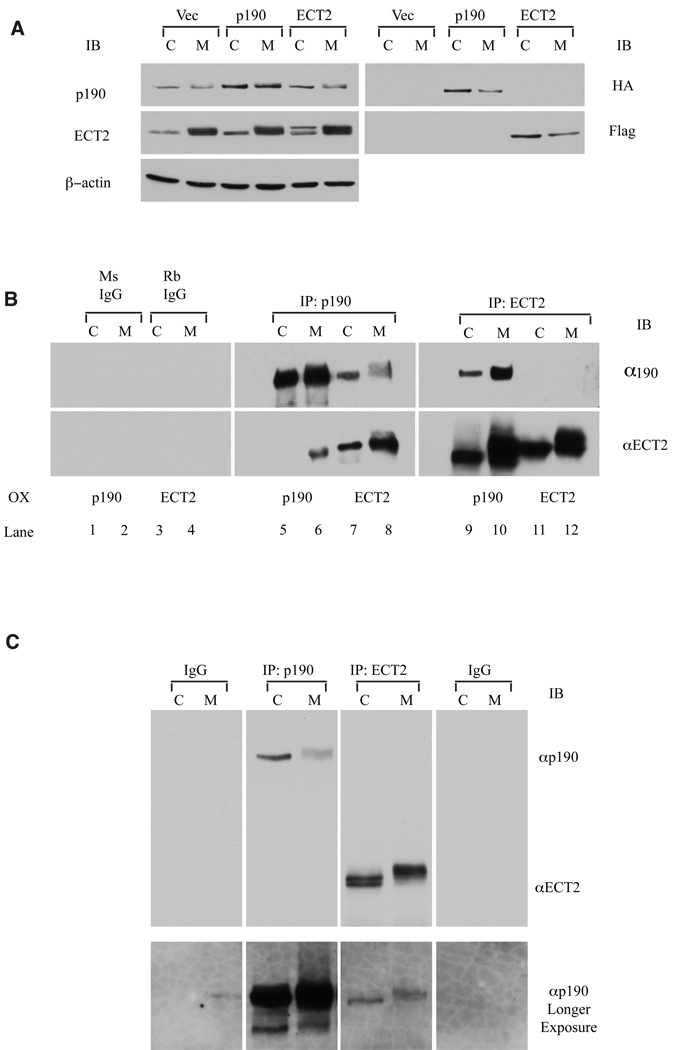
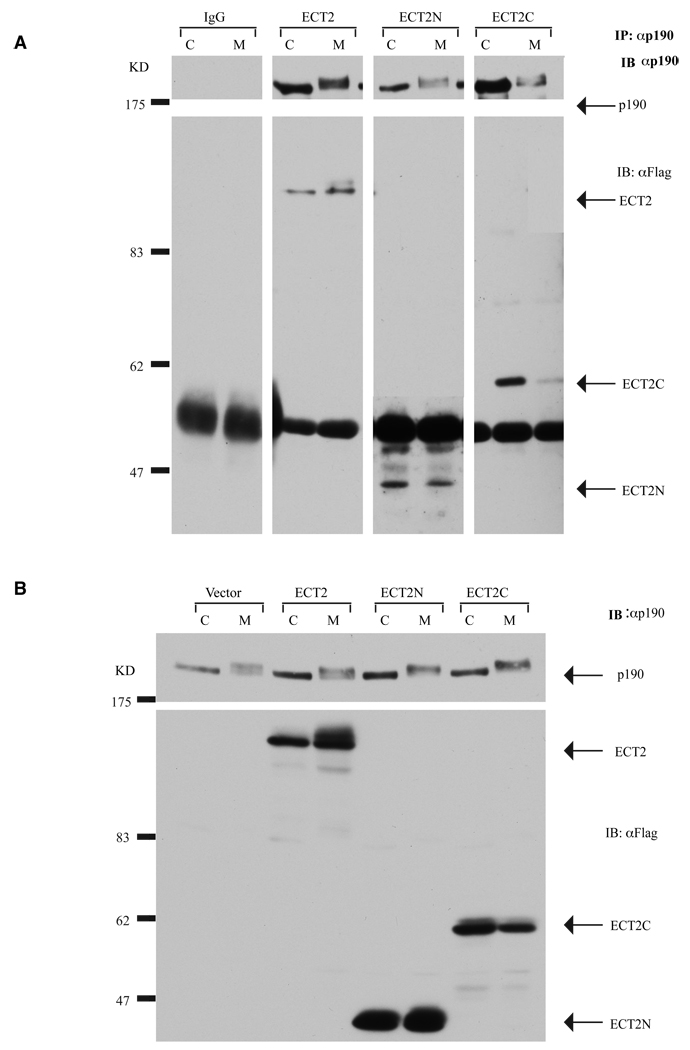
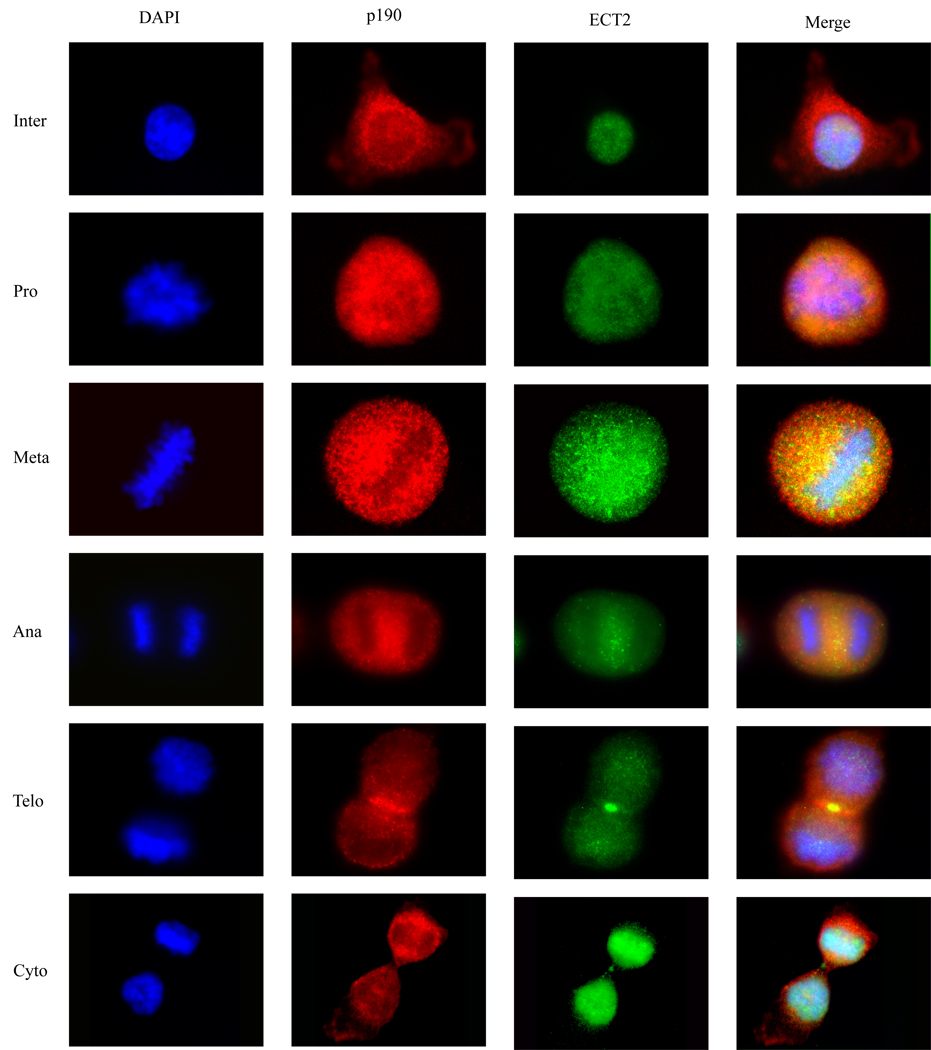
Evidence suggests that p190RhoGAP (p190), a GTPase activating protein (GAP) specific for Rho, plays a role in cytokinesis. First, ectopic expression of p190 induces a multinucleated cellular phenotype. Second, endogenous p190 localizes to the cleavage furrow of dividing cells. Lastly, its levels are reduced in late mitosis by ubiquitin-mediated proteasomal degradation, consistent with the idea that low levels of p190 and high levels of active Rho are required for completion of cytokinesis. As with p190, RhoA and the RhoGEF, ECT2, have been localized to the cleavage furrow. These findings raise the question of whether p190 and ECT2 cooperate antagonistically to regulate the activity of Rho and contraction of the actomyosin ring during cytokinesis. Here we demonstrate ECT2 can, in a dose-dependent manner, reduce multinucleation induced by p190. Furthermore, endogenous p190 and ECT2 colocalize at the cleavage furrow of dividing cells and stably associate with one another in co-immunoprecipitation assays. Functional and physical interactions between p190 and ECT2 are reflected in the levels of Rho activity, as assessed by Rho pull-down assays. Together, these results suggest that co-regulation of Rho activity by p190RhoGAP and ECT2 in the cleavage furrow determines whether cells properly complete cytokinesis.
Figures

References
-
- Brouns MR, Matheson SF, Hu KQ, Delalle I, Caviness VS, Silver J, Bronson RT, Settleman J. The adhesion signaling molecule p190 RhoGAP is required for morphogenetic processes in neural development. Development. 2000;127:4891–4903. - PubMed
-
- Brouns MR, Matheson SF, Settleman J. p190 RhoGAP is the principal Src substrate in brain and regulates axon outgrowth, guidance and fasciculation. Nat. Cell Biol. 2001;3:361–367. - PubMed
-
- Chalamalasetty RB, Hummer S, Nigg EA, Sillje HH. Influence of human Ect2 depletion and overexpression on cleavage furrow formation and abscission. J. Cell Sci. 2006;119:3008–3019. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous