

Staphylococcus aureus haem oxygenases are differentially regulated by iron and haem
- PMID: 18643935
- PMCID: PMC2597461
- DOI: 10.1111/j.1365-2958.2008.06363.x
Staphylococcus aureus haem oxygenases are differentially regulated by iron and haem
Abstract
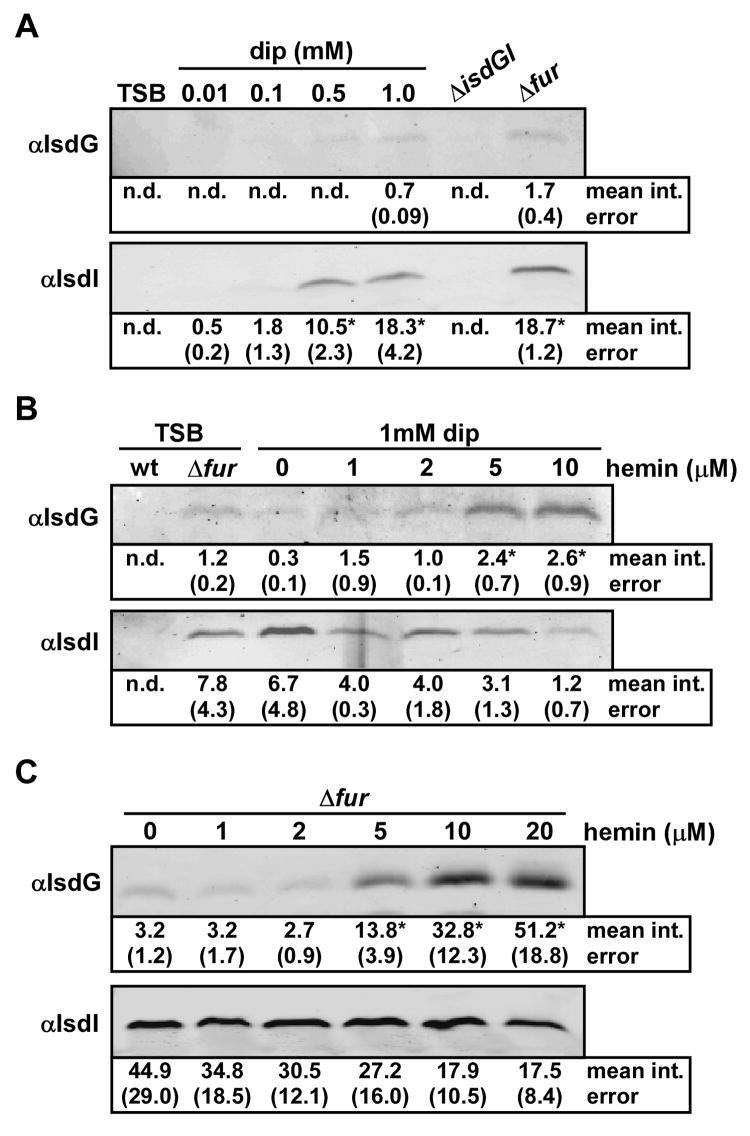
Iron acquisition is a vital process for most pathogenic bacteria, as iron is a limiting nutrient during infection. Staphylococcus aureus, an increasingly important pathogen, acquires iron from host haem via elaboration of the iron-regulated surface determinant system (Isd). IsdG and IsdI are haem oxygenases that have been proposed to degrade exogenous haem in the bacterial cytoplasm as a mechanism to liberate free iron for use as a nutrient source. Herein, we report that IsdG and IsdI are both important for S. aureus growth on haemin as a sole iron source and are necessary for full S. aureus pathogenesis. Investigations into the regulation of these enzymes revealed that IsdG and IsdI are differentially regulated by iron and haem through both transcriptional and post-transcriptional mechanisms. Additionally, IsdI was found to be expressed in infected tissues at the sites of abscess formation, suggesting that abscesses are iron-starved microenvironments inside the host. These findings suggest that S. aureus differentially regulates IsdG and IsdI in response to alterations in iron and haem availability during infection.
Figures






References
-
- Bae T, Schneewind O. Allelic replacement in Staphylococcus aureus with inducible counter-selection. Plasmid. 2006;55:58–63. - PubMed
-
- Braun V, Gunter K, Hantke K. Transport of iron across the outer membrane. Biol Met. 1991;4:14–22. - PubMed
-
- Bullen JJ, Griffiths E. Iron and Infection: Molecular, Physiological and Clinical Aspects. New York: John Wiley and Sons; 1999.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
