

Review
doi: 10.1016/j.mib.2008.06.004.
Epub 2008 Jul 31.
Structural proteomics of dengue virus
Affiliations
- PMID: 18644250
- PMCID: PMC2581888
- DOI: 10.1016/j.mib.2008.06.004
Item in Clipboard
Review
Structural proteomics of dengue virus
Curr Opin Microbiol.
2008 Aug.
Abstract
In this paper, we discuss recent advances in our knowledge of the dengue virus life cycle based on new structural data of the virus and its proteins. Specifically, we focus on the structure of the pre-membrane protein, prM and its role in virus assembly, the first full-length structure of a multi-domain dengue virus replication protein, NS3, and the recently solved structures of NS5 methyltransferase and polymerase domains. These structures provide a basis for describing function and predicting putative host interactions.
Figures
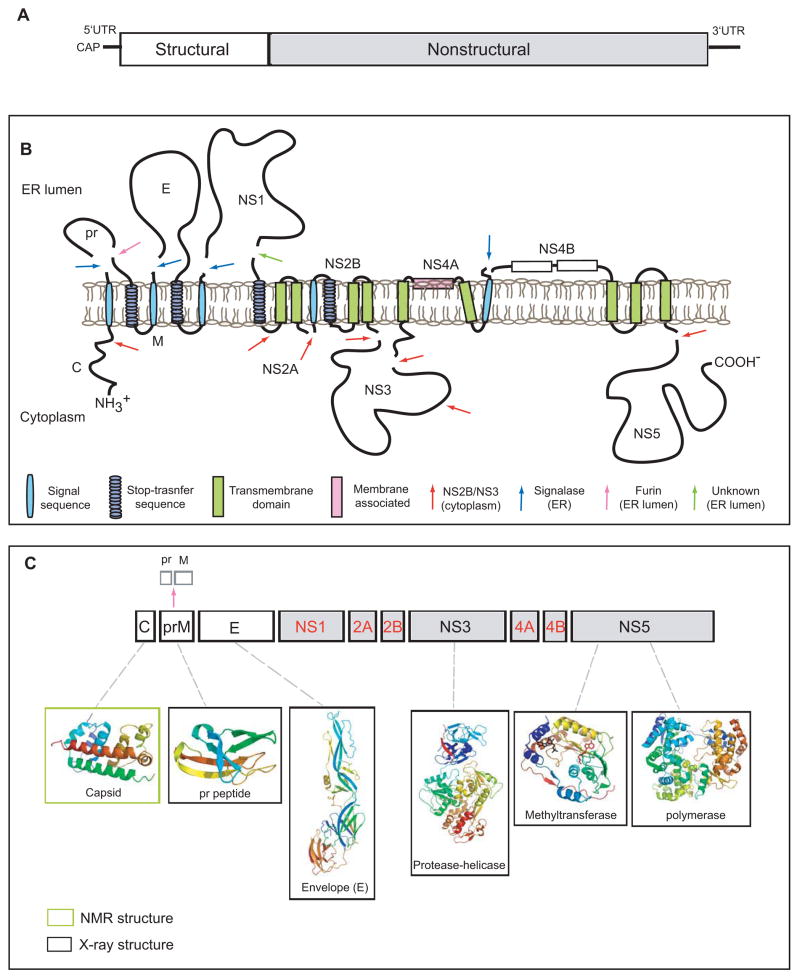
A. The viral genome is a positive sense RNA of ~11kb in length. It is capped at the 5′ end but lacks a poly-(A) at its 3′-end. The structural proteins (open boxes) are encoded at the 5′ one third of genome followed by the non-structural proteins (grey boxes). B. Membrane topology of the polyprotein. The viral RNA is translated as a polyprotein and processed by cellular and viral proteases (denoted by arrows). The structural proteins include capsid (C), membrane protein (prM/M) and envelope (E). prM and E are released from the polyprotein by signalase cleavage in the ER, but remain anchored on the luminal side of the membrane. The C is also anchored in the ER membrane (on the cytoplasmic side) by a conserved hydrophobic signal sequence at its C-terminal end. This signal sequence is cleaved by the viral NS2B-NS3 protease. During virus maturation, prM is further cleaved by furin in the TGN into the pr peptide and M protein. The non-structural proteins are processed mainly by the viral protease NS2B-NS3 in the cytoplasm with the exception of NS1, which is released from NS2A by a yet unidentified protease in the lumen of the ER. NS2A/2B and NS4A/4B are anchored in the ER as transmembrane proteins. The topology of NS4A and NS4B are predicted through biochemical and cellular analyses [62,67]. C. Structural proteome of dengue virus. NMR and X-ray structures are shown for C, prM, E, NS3 (full-length) and the NS5 methyltransferase and polymerase domains (PDB identifiers: 1R6R, 3C5X, 1OKE, 2VBC, 2P1D, 2J7U, respectively)[, –7, 12, 13, 18, 32, 36]. Structures are currently not available for the proteins denoted in red.
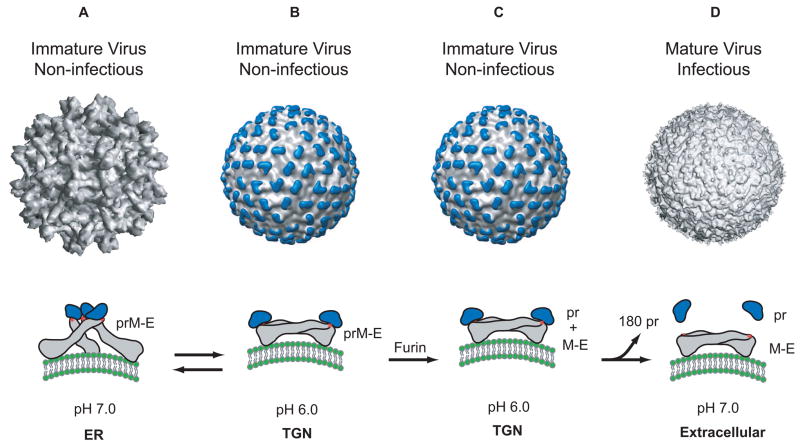
Structure of the dengue virion and conformations of the E protein. A. The cryo-EM reconstruction of the immature virion at neutral pH [12]. In this structure, the E protein exists as a heterodimer with prM, and these heterodimers form 60 trimeric spikes that extend away from the surface of the virus. This arrangement of E gives the virus a ‘spiky’ morphology and represents the initial particle that buds into the ER. The conformation of the E protein (grey) within a spike is shown below the virion. The ‘pr’ peptide is shown in blue protecting the fusion peptide on E (shown as a red star). B. The cryo-EM reconstruction of the immature virion at low pH [14]. During its transit through the secretory pathway, the virus encounters low pH in the TGN. Under these conditions, the prM-E heterodimers dissociate from their trimeric spike-like organization and form 90 dimers that lie flat against the viral surface. This orientation of prM-E proteins gives the virion a ‘smooth’ morphology. Upon raising the pH (reverse reaction), this ‘smooth’ particle can revert back to its ‘spiky’ morphology. C. While in the TGN, the prM protein is cleaved into its ‘pr’ peptide and M protein by the host endoprotease, furin. The cleaved ‘pr’ peptide maintains its position as a ‘cap’ on E and the E proteins remain as 90 homodimers lying parallel to the virion surface. M, not shown in this figure lies embedded in the viral membrane beneath the E protein shell. D. The cryo-EM reconstruction of the mature virion [10]. Following furin cleavage, the mature virion is secreted into the extracellular milleu and the pr peptide is released from mature particle.
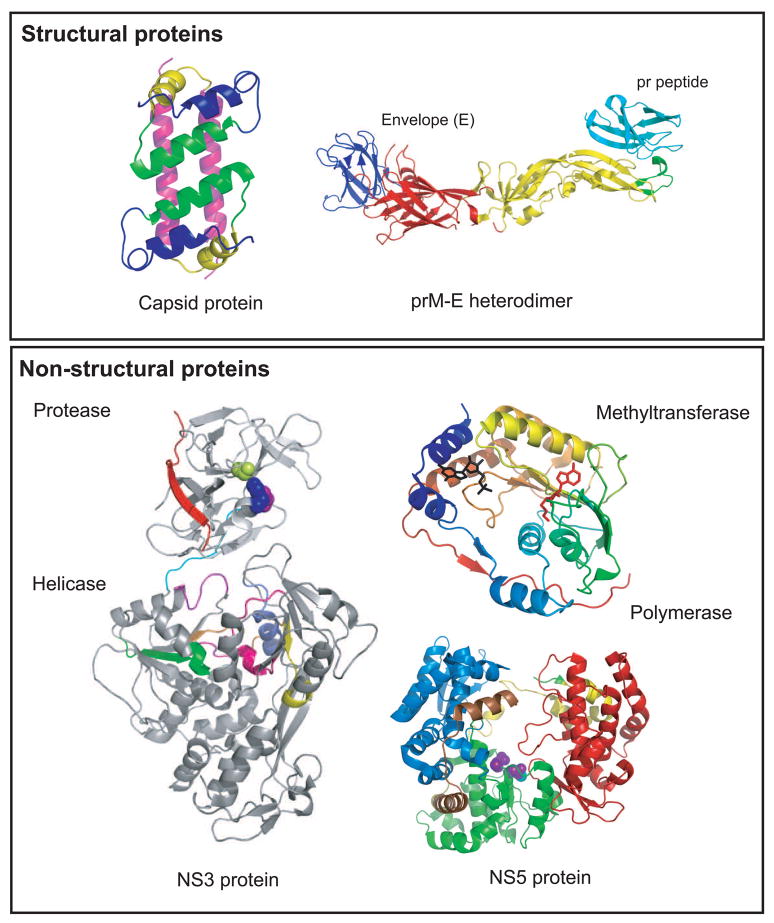
Structures of dengue virus proteins. Top panel (structural proteins): The NMR structure of the capsid protein dimer (1R6R) is shown on the left with helix 1, blue; helix 2, green, helix 3, yellow and helix 4, pink [4]. The X-ray structure of the prM-E protein is shown on the right [13]. The three domains of the E protein are shown: domain I, red; domain II, yellow; domain III, blue. prM is shown in cyan forming a cap protecting the fusion peptide (green) on E. Bottom panel (non-structural proteins): the X-ray structure of the full-length NS3 protein (2VBC) is shown on the left [18]. The protease domain (residues 1–168) is N-terminally located in the molecule. The catalytic triad is shown in blue (His 51), purple (Asp 75) and limon (Ser 135). NS2B is shown in red. The linker between the protease and helicase domains (residues 169–179) is shown in cyan. The helicase domain forms the C-terminal two-thirds of the molecule (residues 180–618). The active site motifs are colored: motif I (196–202), purple; motif II (284–287), magenta; motif III (312–324), orange; motif IV (359–367), yellow; motif V (407–420), pink; motif VI (452–463), slate blue. NS5 is shown on the right. The methytransferase (2P1D) is shown colored in rainbow from N-terminal to the C-terminal end [32]. The CAP structure is shown in black and the by-product of the methylation reaction, S-adenosyl-homocysteine (SAH) is shown in red. The polymerase of DENV-3 (2J7U) is shown with the fingers domain (273–315; 416–496; 543–600) in blue, palm domain (497–542; 601–705) in green, and thumb domain (706–900) in red. The nuclear localization signal (NLS) is shown in yellow (b NLS, 320–368) and brown (a/b NLS, 369–405).
References
-
- Morens DM, Fauci AS. Dengue and hemorrhagic fever: a potential threat to public health in the United States. Jama. 2008;299:214–216. - PubMed
-
- Gould EA, Solomon T. Pathogenic flaviviruses. Lancet. 2008;371:500–509. - PubMed
-
- Kuhn RJ. The Flaviviruses. In: Acheson NH, editor. Fundamentals of Molecular Virology. 2004. in press.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
