

Ascidians and the plasticity of the chordate developmental program
- PMID: 18644342
- PMCID: PMC2922861
- DOI: 10.1016/j.cub.2008.05.039
Ascidians and the plasticity of the chordate developmental program
Abstract
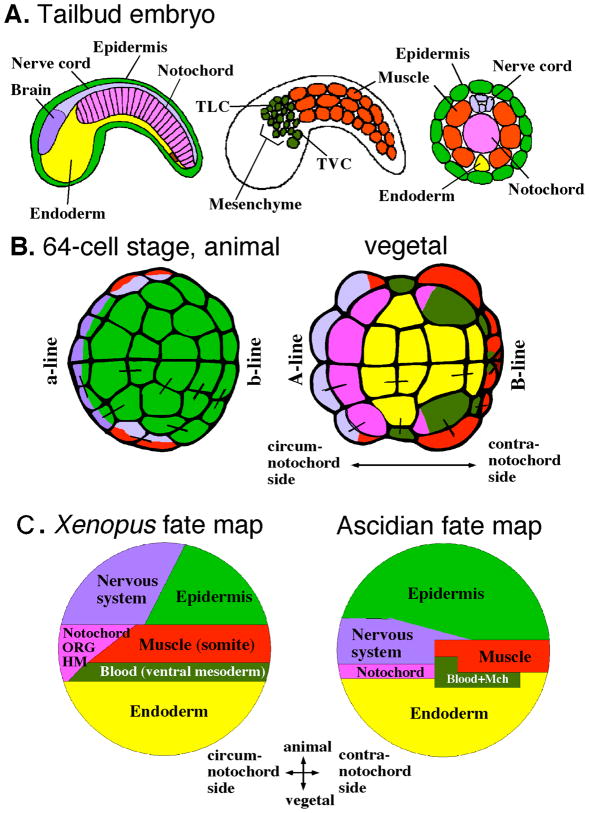
Little is known about the ancient chordates that gave rise to the first vertebrates, but the descendants of other invertebrate chordates extant at the time still flourish in the ocean. These invertebrates include the cephalochordates and tunicates, whose larvae share with vertebrate embryos a common body plan with a central notochord and a dorsal nerve cord. Tunicates are now thought to be the sister group of vertebrates. However, research based on several species of ascidians, a diverse and wide-spread class of tunicates, revealed that the molecular strategies underlying their development appear to diverge greatly from those found in vertebrates. Furthermore, the adult body plan of most tunicates, which arises following an extensive post-larval metamorphosis, shows little resemblance to the body plan of any other chordate. In this review, we compare the developmental strategies of ascidians and vertebrates and argue that the very divergence of these strategies reveals the surprising level of plasticity of the chordate developmental program and is a rich resource to identify core regulatory mechanisms that are evolutionarily conserved in chordates. Further, we propose that the comparative analysis of the architecture of ascidian and vertebrate gene regulatory networks may provide critical insight into the origin of the chordate body plan.
Figures





References
-
- Haeckel E. Die Gastrea-Theorie, die phylogenetische Klassification des Tierreiches und Homologie der Keimblätter. Jena Z Naturwiss. 1874;8:1–55.
-
- Alldredge AaLPM. Pelagic tunicates: unique herbivores in the marine plankton. Bioscience. 1982;32:655–663.
-
- Satoh N. Developmental Biology of Ascidians. Cambridge: Cambridge University Press; 1994.
-
- Delsuc F, Brinkmann H, Chourrout D, Philippe H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature. 2006;439:965–968. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
