

Hormone-dependent nuclear export of estradiol receptor and DNA synthesis in breast cancer cells
- PMID: 18644889
- PMCID: PMC2483513
- DOI: 10.1083/jcb.200712125
Hormone-dependent nuclear export of estradiol receptor and DNA synthesis in breast cancer cells
Abstract
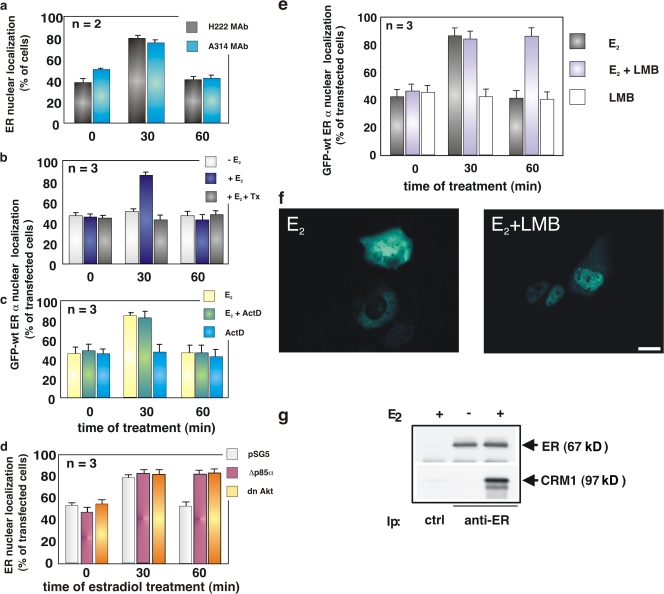
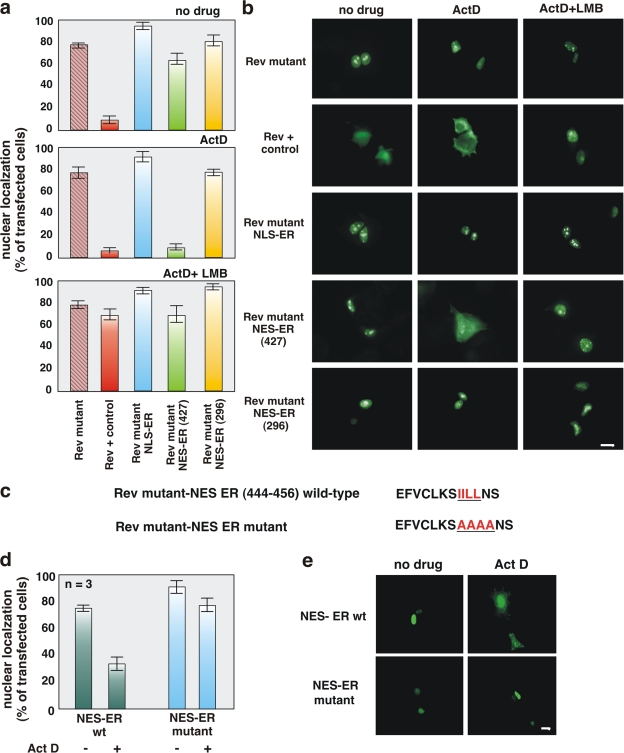
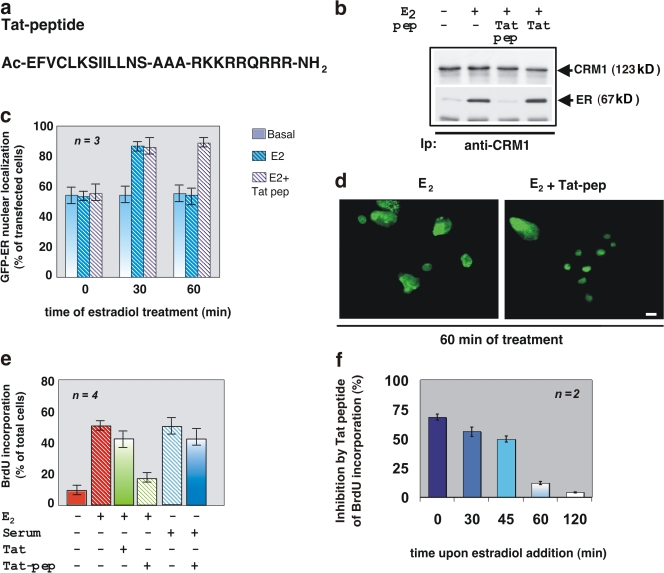
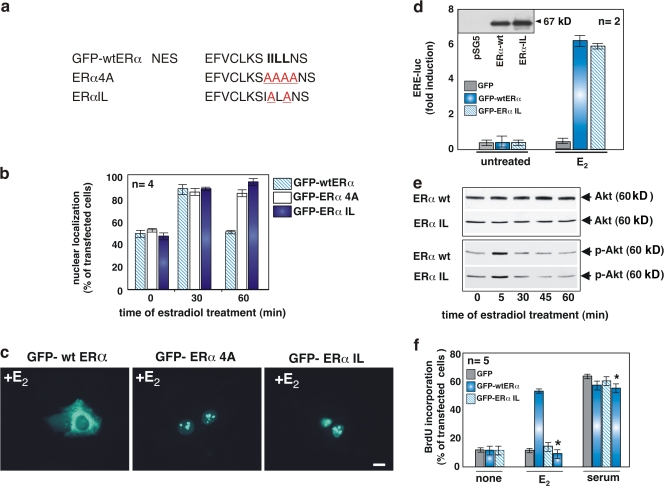
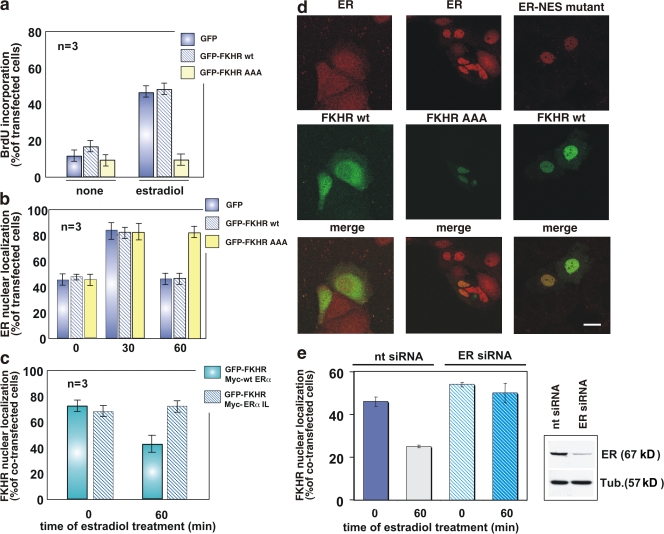
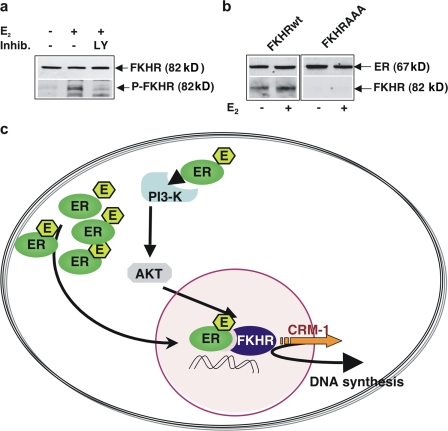
In breast cancer cells, cytoplasmic localization of the estradiol receptor alpha (ERalpha) regulates estradiol-dependent S phase entry. We identified a nuclear export sequence (NES) in ERalpha and show that its export is dependent on both estradiol-mediated phosphatidylinositol-3-kinase (PI3K)/AKT activation and chromosome region maintenance 1 (CRM1). A Tat peptide containing the ERalpha NES disrupts ERalpha-CRM1 interaction and prevents nuclear export of ERalpha- and estradiol-induced DNA synthesis. NES-ERalpha mutants do not exit the nucleus and inhibit estradiol-induced S phase entry; ERalpha-dependent transcription is normal. ERalpha is associated with Forkhead proteins in the nucleus, and estradiol stimulates nuclear exit of both proteins. ERalpha knockdown or ERalpha NES mutations prevent ERalpha and Forkhead nuclear export. A mutant of forkhead in rhabdomyosarcoma (FKHR), which cannot be phosphorylated by estradiol-activated AKT, does not associate with ERalpha and is trapped in the nucleus, blocking S phase entry. In conclusion, estradiol-induced AKT-dependent phosphorylation of FKHR drives its association with ERalpha, thereby triggering complex export from the nucleus necessary for initiation of DNA synthesis and S phase entry.
Figures
References
-
- Birkenkamp, K.U., and P.J. Coffer. 2003. Regulation of cell survival and proliferation by the FOXO (Forkhead box, class O) subfamily of Forkhead transcription factors. Biochem. Soc. Trans. 31:292–297. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
