

Crenarchaeal arginine decarboxylase evolved from an S-adenosylmethionine decarboxylase enzyme
- PMID: 18650422
- PMCID: PMC2533785
- DOI: 10.1074/jbc.M802674200
Crenarchaeal arginine decarboxylase evolved from an S-adenosylmethionine decarboxylase enzyme
Abstract
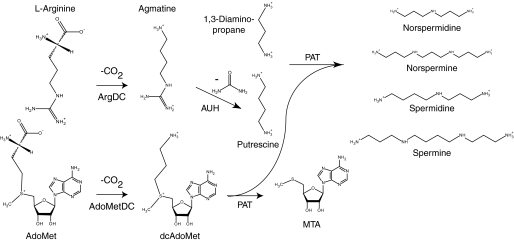
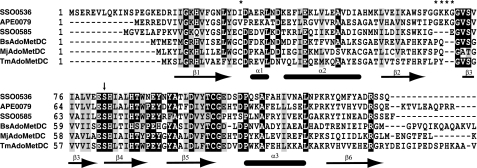
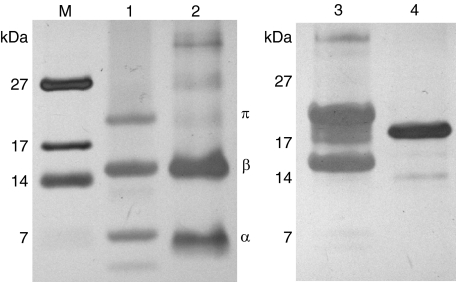
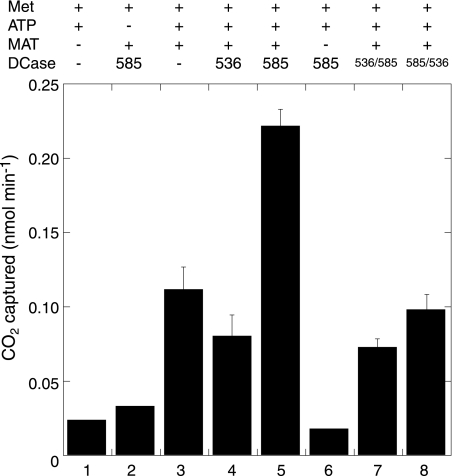
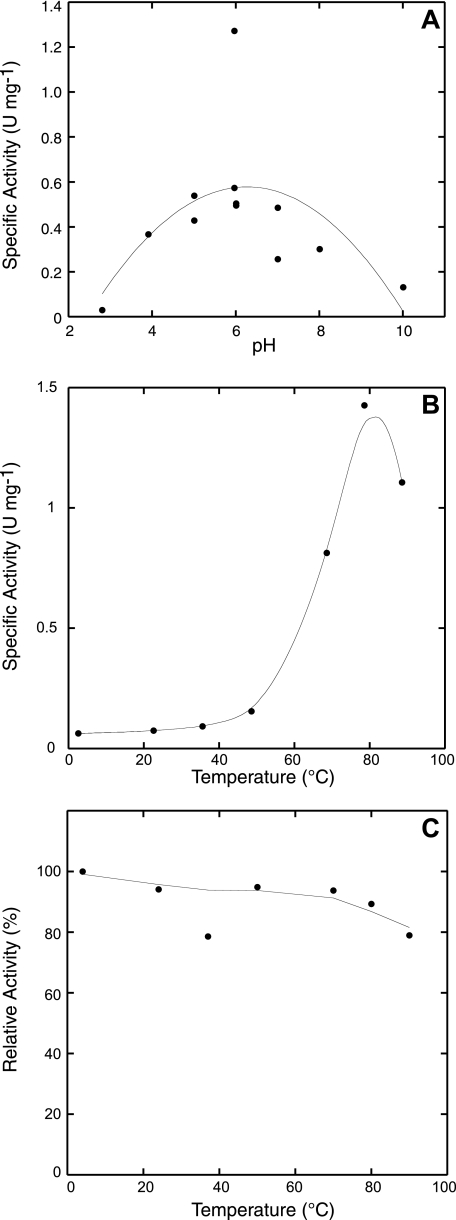
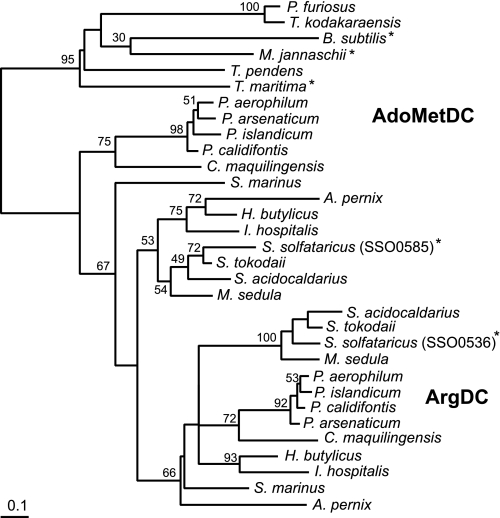
The crenarchaeon Sulfolobus solfataricus uses arginine to produce putrescine for polyamine biosynthesis. However, genome sequences from S. solfataricus and most crenarchaea have no known homologs of the previously characterized pyridoxal 5'-phosphate or pyruvoyl-dependent arginine decarboxylases that catalyze the first step in this pathway. Instead they have two paralogs of the S-adenosylmethionine decarboxylase (AdoMetDC). The gene at locus SSO0585 produces an AdoMetDC enzyme, whereas the gene at locus SSO0536 produces a novel arginine decarboxylase (ArgDC). Both thermostable enzymes self-cleave at conserved serine residues to form amino-terminal beta-domains and carboxyl-terminal alpha-domains with reactive pyruvoyl cofactors. The ArgDC enzyme specifically catalyzed arginine decarboxylation more efficiently than previously studied pyruvoyl enzymes. alpha-Difluoromethylarginine significantly reduced the ArgDC activity of purified enzyme, and treating growing S. solfataricus cells with this inhibitor reduced the cells' ratio of spermidine to norspermine by decreasing the putrescine pool. The crenarchaeal ArgDC had no AdoMetDC activity, whereas its AdoMetDC paralog had no ArgDC activity. A chimeric protein containing the beta-subunit of SSO0536 and the alpha-subunit of SSO0585 had ArgDC activity, implicating residues responsible for substrate specificity in the amino-terminal domain. This crenarchaeal ArgDC is the first example of alternative substrate specificity in the AdoMetDC family. ArgDC activity has evolved through convergent evolution at least five times, demonstrating the utility of this enzyme and the plasticity of amino acid decarboxylases.
Figures
Similar articles
-
Neofunctionalization of S-adenosylmethionine decarboxylase into pyruvoyl-dependent L-ornithine and L-arginine decarboxylases is widespread in bacteria and archaea.J Biol Chem. 2023 Aug;299(8):105005. doi: 10.1016/j.jbc.2023.105005. Epub 2023 Jul 1. J Biol Chem. 2023. PMID: 37399976 Free PMC article.
-
Methanococcus jannaschii uses a pyruvoyl-dependent arginine decarboxylase in polyamine biosynthesis.J Biol Chem. 2002 Jun 28;277(26):23500-7. doi: 10.1074/jbc.M203467200. Epub 2002 Apr 29. J Biol Chem. 2002. PMID: 11980912
-
Trypanosoma cruzi has not lost its S-adenosylmethionine decarboxylase: characterization of the gene and the encoded enzyme.Biochem J. 1998 Aug 1;333 ( Pt 3)(Pt 3):527-37. doi: 10.1042/bj3330527. Biochem J. 1998. PMID: 9677309 Free PMC article.
-
Structural biology of S-adenosylmethionine decarboxylase.Amino Acids. 2010 Feb;38(2):451-60. doi: 10.1007/s00726-009-0404-y. Epub 2009 Dec 8. Amino Acids. 2010. PMID: 19997761 Free PMC article. Review.
-
Structural and mechanistic properties of E. coli adenosylmethionine decarboxylase.Adv Exp Med Biol. 1988;250:81-9. doi: 10.1007/978-1-4684-5637-0_8. Adv Exp Med Biol. 1988. PMID: 3076348 Review.
Cited by
-
Augmenting the genetic toolbox for Sulfolobus islandicus with a stringent positive selectable marker for agmatine prototrophy.Appl Environ Microbiol. 2013 Sep;79(18):5539-49. doi: 10.1128/AEM.01608-13. Epub 2013 Jul 8. Appl Environ Microbiol. 2013. PMID: 23835176 Free PMC article.
-
Functional polyamine metabolic enzymes and pathways encoded by the virosphere.Proc Natl Acad Sci U S A. 2023 Feb 28;120(9):e2214165120. doi: 10.1073/pnas.2214165120. Epub 2023 Feb 21. Proc Natl Acad Sci U S A. 2023. PMID: 36802435 Free PMC article.
-
Candidatus Nitrosocaldus cavascurensis, an Ammonia Oxidizing, Extremely Thermophilic Archaeon with a Highly Mobile Genome.Front Microbiol. 2018 Jan 26;9:28. doi: 10.3389/fmicb.2018.00028. eCollection 2018. Front Microbiol. 2018. PMID: 29434576 Free PMC article.
-
Development of the Multiple Gene Knockout System with One-Step PCR in Thermoacidophilic Crenarchaeon Sulfolobus acidocaldarius.Archaea. 2017 Oct 31;2017:7459310. doi: 10.1155/2017/7459310. eCollection 2017. Archaea. 2017. PMID: 29225512 Free PMC article.
-
Prediction and Analysis of Post-Translational Pyruvoyl Residue Modification Sites from Internal Serines in Proteins.PLoS One. 2013 Jun 21;8(6):e66678. doi: 10.1371/journal.pone.0066678. Print 2013. PLoS One. 2013. PMID: 23805260 Free PMC article.
References
-
- De Rosa, M., De Rosa, S., Gambacorta, A., Cartenì-Farina, M., and Zappia, V. (1976) Biochem. Biophys. Res. Commun. 69 253–261 - PubMed
-
- Hamana, K., Hamana, H., Niitsu, M., Samejima, K., Sakane, T., and Yokota, A. (1994) Microbios 79 109–119 - PubMed
-
- Cacciapuoti, G., Porcelli, M., De Rosa, M., Gambacorta, A., Bertoldo, C., and Zappia, V. (1991) Eur. J. Biochem. 199 395–400 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
