

Regulatory ATPase sites of cytoplasmic dynein affect processivity and force generation
- PMID: 18650442
- PMCID: PMC2533788
- DOI: 10.1074/jbc.M802951200
Regulatory ATPase sites of cytoplasmic dynein affect processivity and force generation
Abstract
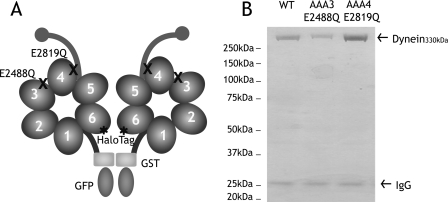
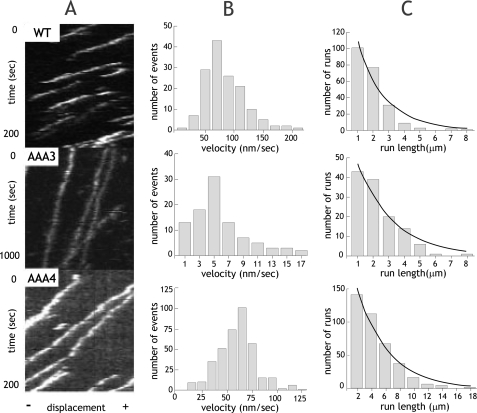
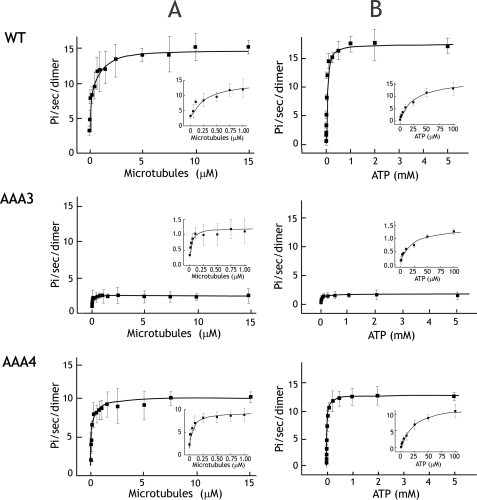
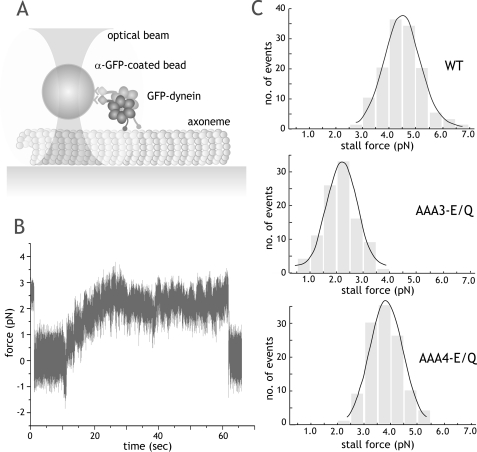
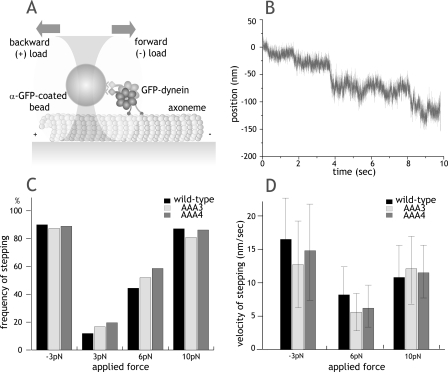
The heavy chain of cytoplasmic dynein contains four nucleotide-binding domains referred to as AAA1-AAA4, with the first domain (AAA1) being the main ATP hydrolytic site. Although previous studies have proposed regulatory roles for AAA3 and AAA4, the role of ATP hydrolysis at these sites remains elusive. Here, we have analyzed the single molecule motility properties of yeast cytoplasmic dynein mutants bearing mutations that prevent ATP hydrolysis at AAA3 or AAA4. Both mutants remain processive, but the AAA4 mutant exhibits a surprising increase in processivity due to its tighter affinity for microtubules. In addition to changes in motility characteristics, AAA3 and AAA4 mutants produce less maximal force than wild-type dynein. These results indicate that the nucleotide binding state at AAA3 and AAA4 can allosterically modulate microtubule binding affinity and affect dynein processivity and force production.
Figures
References
-
- Vallee, R. B., Williams, J. C., Varma, D., and Barnhart, L. E. (2004) J. Neurobiol. 58 189–200 - PubMed
-
- Ogura, T., and Wilkinson, A. J. (2001) Genes Cells 6 575–597 - PubMed
-
- Neuwald, A. F., Aravind, L., Spouge, J. L., and Koonin, E. V. (1999) Genome Res. 9 27–43 - PubMed
-
- Erzberger, J. P., and Berger, J. M. (2006) Annu. Rev. Biophys. Biomol. Struct. 35 93–114 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
