

Cell-specific ATP7A transport sustains copper-dependent tyrosinase activity in melanosomes
- PMID: 18650808
- PMCID: PMC2812007
- DOI: 10.1038/nature07163
Cell-specific ATP7A transport sustains copper-dependent tyrosinase activity in melanosomes
Abstract
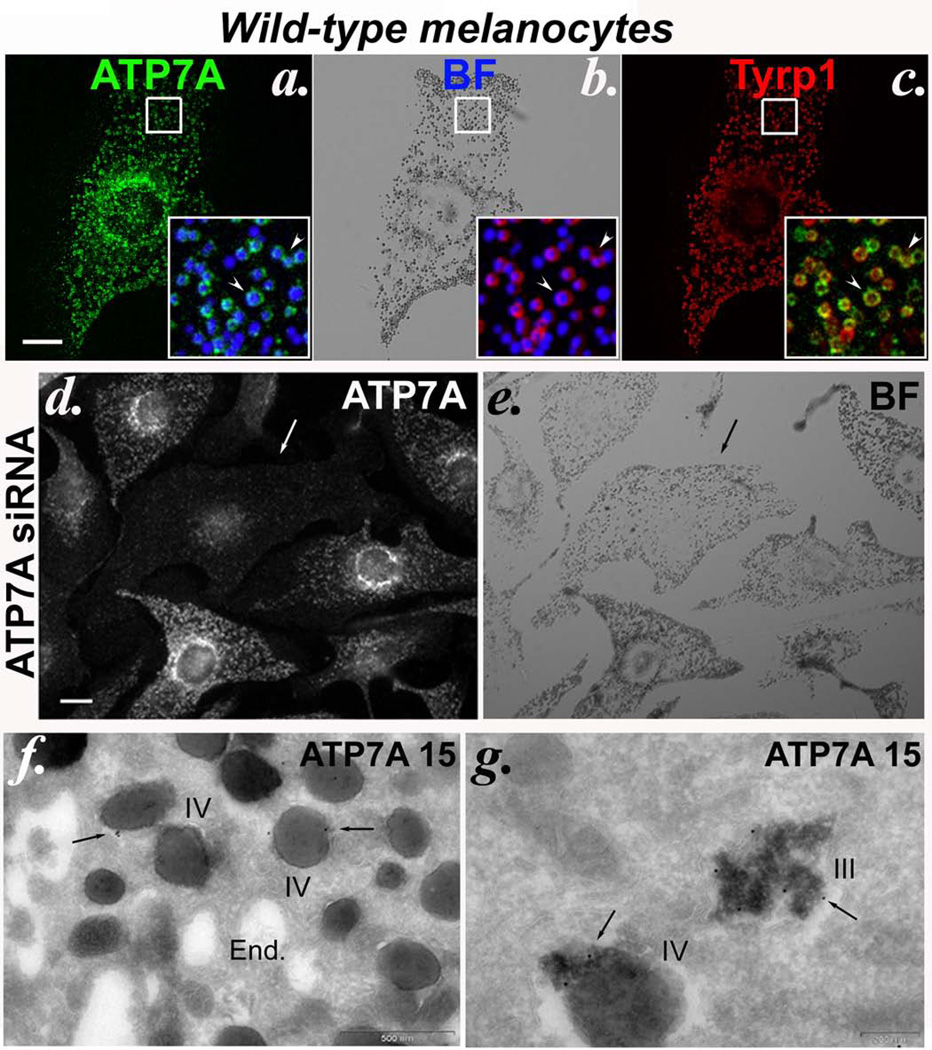
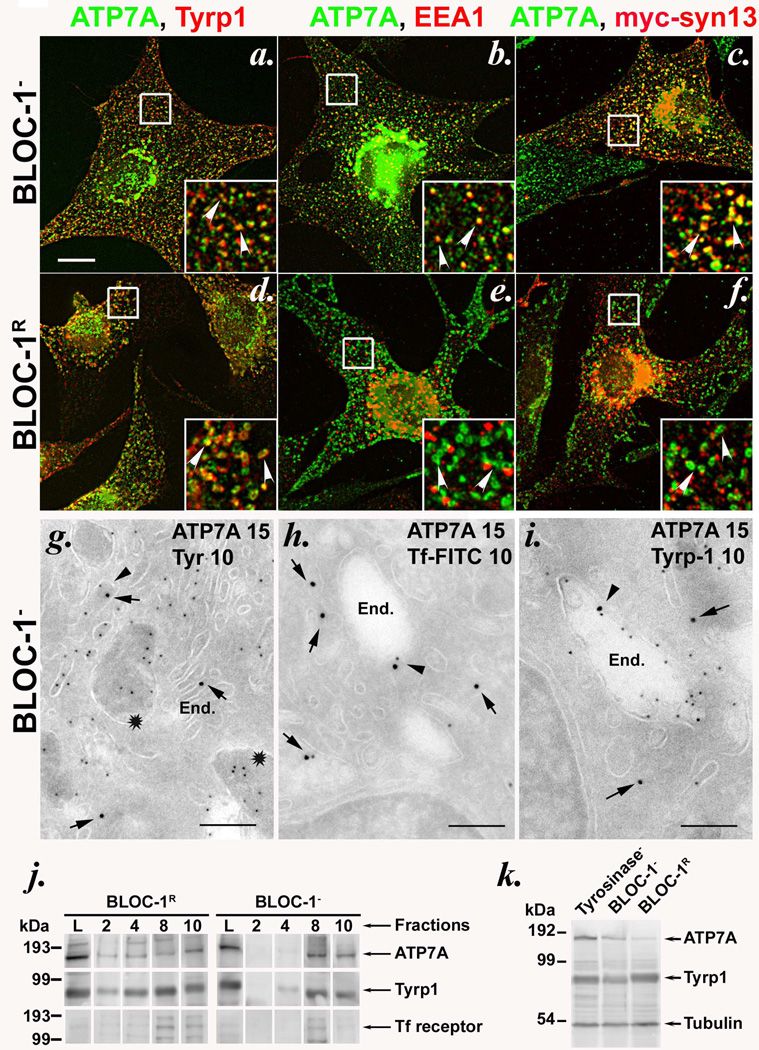
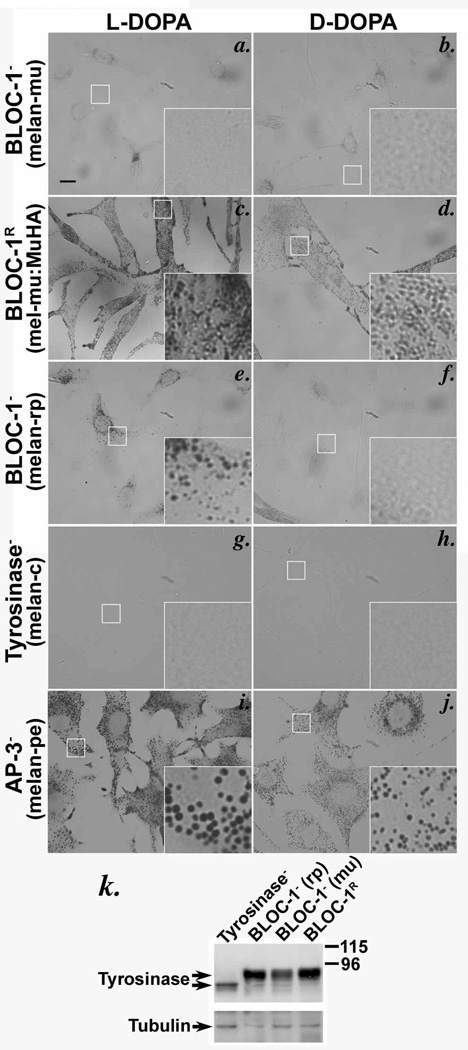
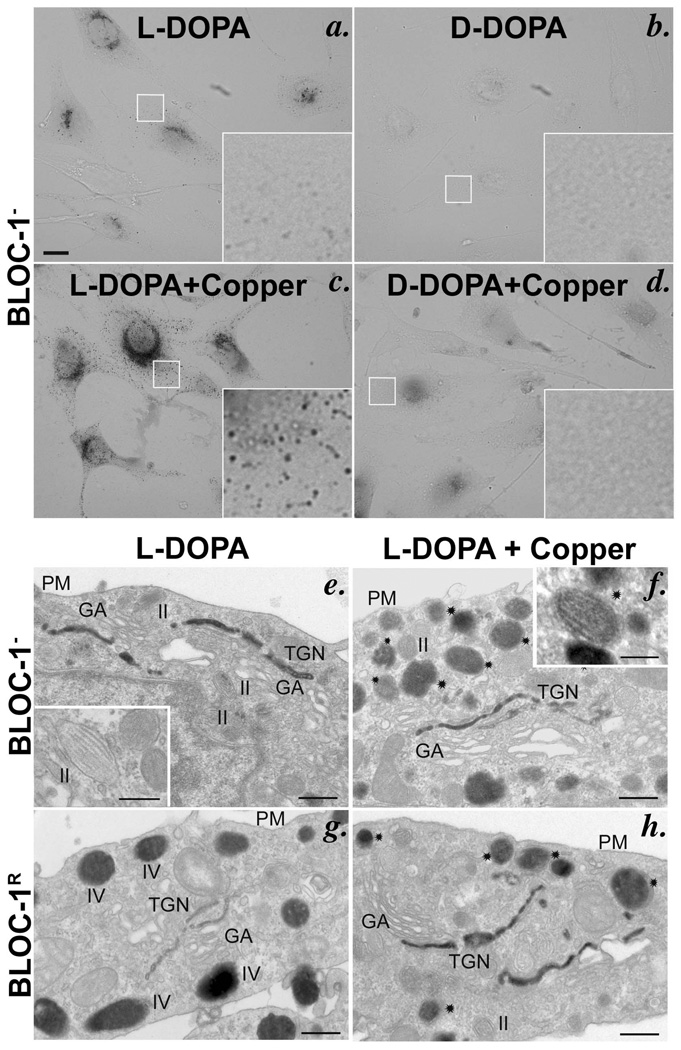
Copper is a cofactor for many cellular enzymes and transporters. It can be loaded onto secreted and endomembrane cuproproteins by translocation from the cytosol into membrane-bound organelles by ATP7A or ATP7B transporters, the genes for which are mutated in the copper imbalance syndromes Menkes disease and Wilson disease, respectively. Endomembrane cuproproteins are thought to incorporate copper stably on transit through the trans-Golgi network, in which ATP7A accumulates by dynamic cycling through early endocytic compartments. Here we show that the pigment-cell-specific cuproenzyme tyrosinase acquires copper only transiently and inefficiently within the trans-Golgi network of mouse melanocytes. To catalyse melanin synthesis, tyrosinase is subsequently reloaded with copper within specialized organelles called melanosomes. Copper is supplied to melanosomes by ATP7A, a cohort of which localizes to melanosomes in a biogenesis of lysosome-related organelles complex-1 (BLOC-1)-dependent manner. These results indicate that cell-type-specific localization of a metal transporter is required to sustain metallation of an endomembrane cuproenzyme, providing a mechanism for exquisite spatial control of metalloenzyme activity. Moreover, because BLOC-1 subunits are mutated in subtypes of the genetic disease Hermansky-Pudlak syndrome, these results also show that defects in copper transporter localization contribute to hypopigmentation, and hence perhaps other systemic defects, in Hermansky-Pudlak syndrome.
Figures
References
-
- Thiele DJ. Integrating trace element metabolism from the cell to the whole organism. J. Nutr. 2003;133:1579S–1580S. - PubMed
-
- Mercer JF. The molecular basis of copper-transport diseases. Trends Mol. Med. 2001;7:64–69. - PubMed
-
- Harris ED. Cellular copper transport and metabolism. Annu. Rev. Nutr. 2000;20:291–310. - PubMed
-
- Lutsenko S, Barnes NL, Bartee MY, et al. Function and regulation of human copper-transporting ATPases. Physiol. Rev. 2007;87:1011–1046. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
