

Cryo-electron microscopy reveals a novel DNA-binding site on the MCM helicase
- PMID: 18650940
- PMCID: PMC2519097
- DOI: 10.1038/emboj.2008.135
Cryo-electron microscopy reveals a novel DNA-binding site on the MCM helicase
Abstract
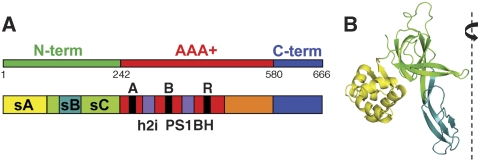
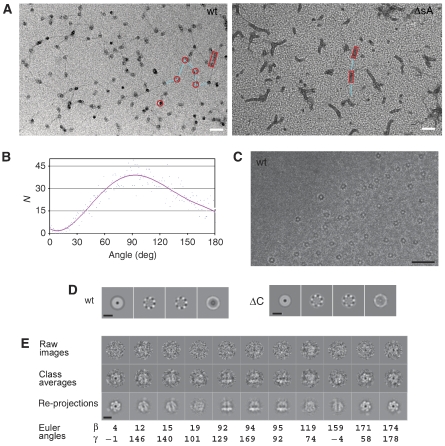
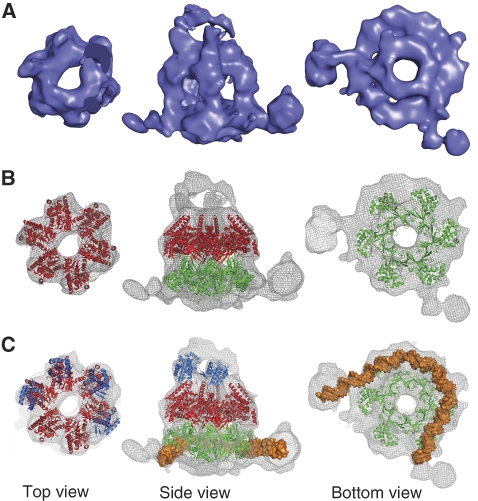
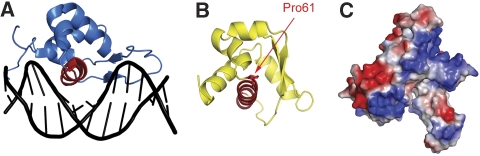
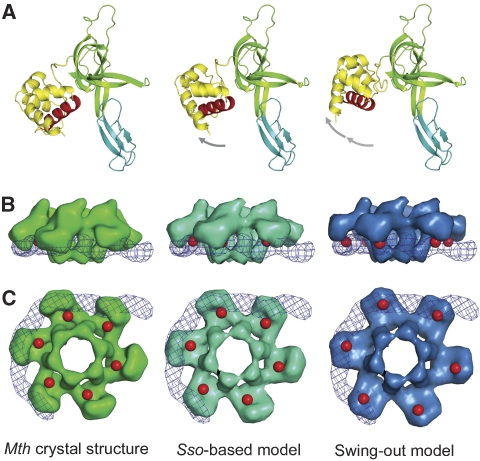
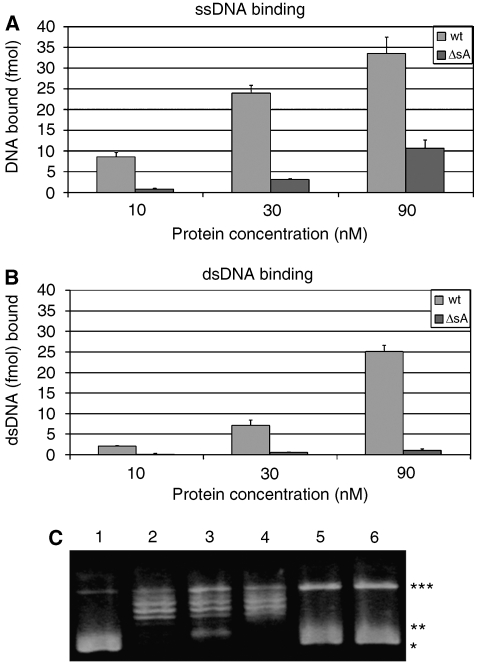
The eukaryotic MCM2-7 complex is recruited at origins of replication during the G1 phase and acts as the main helicase at the replication fork during the S phase of the cell cycle. To characterize the interplay between the MCM helicase and DNA prior to the melting of the double helix, we determined the structure of an archaeal MCM orthologue bound to a 5.6-kb double-stranded DNA segment, using cryo-electron microscopy. DNA wraps around the N-terminal face of a single hexameric ring. This interaction requires a conformational change within the outer belt of the MCM N-terminal domain, exposing a previously unrecognized helix-turn-helix DNA-binding motif. Our findings provide novel insights into the role of the MCM complex during the initiation step of DNA replication.
Figures
References
-
- Aravind L, Anantharaman V, Balaji S, Babu MM, Iyer LM (2005) The many faces of the helix-turn-helix domain: transcription regulation and beyond. FEMS Microbiol Rev 29: 231–262 - PubMed
-
- Arias EE, Walter JC (2007) Strength in numbers: preventing rereplication via multiple mechanisms in eukaryotic cells. Genes Dev 21: 497–518 - PubMed
-
- Boekema EJ, Berden JA, van Heel MG (1986) Structure of mitochondrial F1-ATPase studied by electron microscopy and image processing. Biochim Biophys Acta 851: 353–360 - PubMed
-
- Boisset N, Penczek PA, Taveay JC, You V, de Haas F, Lamy J (1998) Overabundant single-particle electron microscope views induce a three-dimensional reconstruction artifact. Ultramicroscopy 74: 201–207
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
