

30 nm chromatin fibre decompaction requires both H4-K16 acetylation and linker histone eviction
- PMID: 18653199
- PMCID: PMC3870898
- DOI: 10.1016/j.jmb.2008.04.050
30 nm chromatin fibre decompaction requires both H4-K16 acetylation and linker histone eviction
Abstract
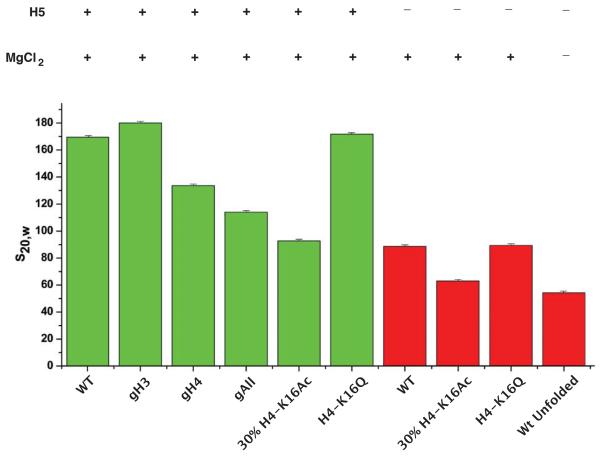
The mechanism by which chromatin is decondensed to permit access to DNA is largely unknown. Here, using a model nucleosome array reconstituted from recombinant histone octamers, we have defined the relative contribution of the individual histone octamer N-terminal tails as well as the effect of a targeted histone tail acetylation on the compaction state of the 30 nm chromatin fiber. This study goes beyond previous studies as it is based on a nucleosome array that is very long (61 nucleosomes) and contains a stoichiometric concentration of bound linker histone, which is essential for the formation of the 30 nm chromatin fiber. We find that compaction is regulated in two steps: Introduction of H4 acetylated to 30% on K16 inhibits compaction to a greater degree than deletion of the H4 N-terminal tail. Further decompaction is achieved by removal of the linker histone.
Figures




Similar articles
-
Histone H4 K16Q mutation, an acetylation mimic, causes structural disorder of its N-terminal basic patch in the nucleosome.J Mol Biol. 2012 Aug 3;421(1):30-7. doi: 10.1016/j.jmb.2012.04.032. Epub 2012 May 7. J Mol Biol. 2012. PMID: 22575889 Free PMC article.
-
ISWI remodelling of physiological chromatin fibres acetylated at lysine 16 of histone H4.PLoS One. 2014 Feb 6;9(2):e88411. doi: 10.1371/journal.pone.0088411. eCollection 2014. PLoS One. 2014. PMID: 24516652 Free PMC article.
-
Regulated acetylation and deacetylation of H4 K16 is essential for efficient NER in Saccharomyces cerevisiae.DNA Repair (Amst). 2018 Dec;72:39-55. doi: 10.1016/j.dnarep.2018.09.009. Epub 2018 Sep 22. DNA Repair (Amst). 2018. PMID: 30274769
-
Secondary structures of the core histone N-terminal tails: their role in regulating chromatin structure.Subcell Biochem. 2013;61:37-55. doi: 10.1007/978-94-007-4525-4_2. Subcell Biochem. 2013. PMID: 23150245 Review.
-
Nuclear matrix, dynamic histone acetylation and transcriptionally active chromatin.Mol Biol Rep. 1997 Aug;24(3):197-207. doi: 10.1023/a:1006811817247. Mol Biol Rep. 1997. PMID: 9291093 Review.
Cited by
-
Dosage compensation in Drosophila melanogaster: epigenetic fine-tuning of chromosome-wide transcription.Nat Rev Genet. 2012 Jan 18;13(2):123-34. doi: 10.1038/nrg3124. Nat Rev Genet. 2012. PMID: 22251873 Review.
-
VRK1 Phosphorylates Tip60/KAT5 and Is Required for H4K16 Acetylation in Response to DNA Damage.Cancers (Basel). 2020 Oct 15;12(10):2986. doi: 10.3390/cancers12102986. Cancers (Basel). 2020. PMID: 33076429 Free PMC article.
-
The tumor suppressor SirT2 regulates cell cycle progression and genome stability by modulating the mitotic deposition of H4K20 methylation.Genes Dev. 2013 Mar 15;27(6):639-53. doi: 10.1101/gad.211342.112. Epub 2013 Mar 6. Genes Dev. 2013. PMID: 23468428 Free PMC article.
-
Epigenetic regulation of epithelial-mesenchymal transition.Cell Mol Life Sci. 2016 Dec;73(23):4493-4515. doi: 10.1007/s00018-016-2303-1. Epub 2016 Jul 8. Cell Mol Life Sci. 2016. PMID: 27392607 Free PMC article. Review.
-
Sex-specific phenotypes of histone H4 point mutants establish dosage compensation as the critical function of H4K16 acetylation in Drosophila.Proc Natl Acad Sci U S A. 2018 Dec 26;115(52):13336-13341. doi: 10.1073/pnas.1817274115. Epub 2018 Dec 10. Proc Natl Acad Sci U S A. 2018. PMID: 30530664 Free PMC article.
References
-
- Kornberg RD. Structure of chromatin. Annu Rev Biochem. 1977;46:931–54. - PubMed
-
- Richmond TJ, Finch JT, Rushton B, Rhodes D, Klug A. Structure of the nucleosome core particle at 7 A resolution. Nature. 1984;311:532–7. - PubMed
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–60. - PubMed
-
- Allan J, Hartman PG, Crane-Robinson C, Aviles FX. The structure of histone H1 and its location in chromatin. Nature. 1980;288:675–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
