

A mitotic GlcNAcylation/phosphorylation signaling complex alters the posttranslational state of the cytoskeletal protein vimentin
- PMID: 18653473
- PMCID: PMC2555957
- DOI: 10.1091/mbc.e07-11-1146
A mitotic GlcNAcylation/phosphorylation signaling complex alters the posttranslational state of the cytoskeletal protein vimentin
Abstract
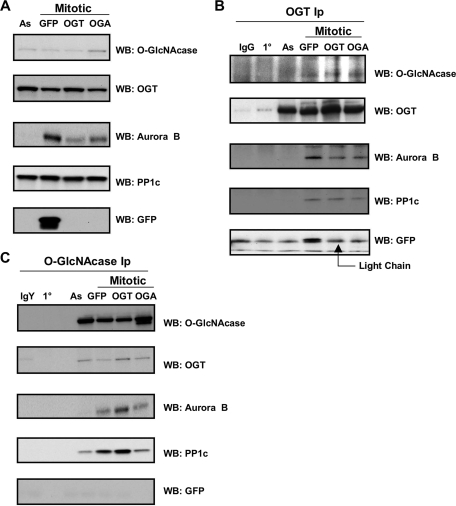
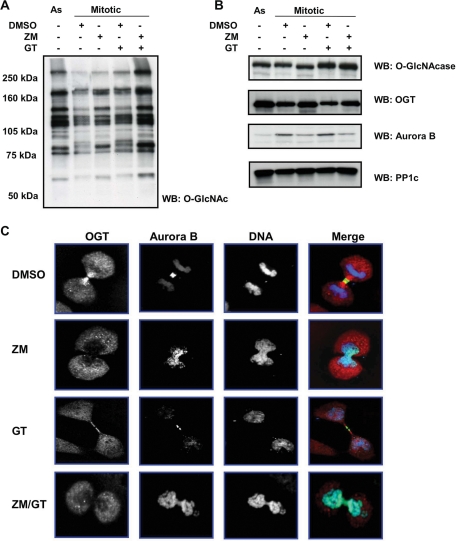
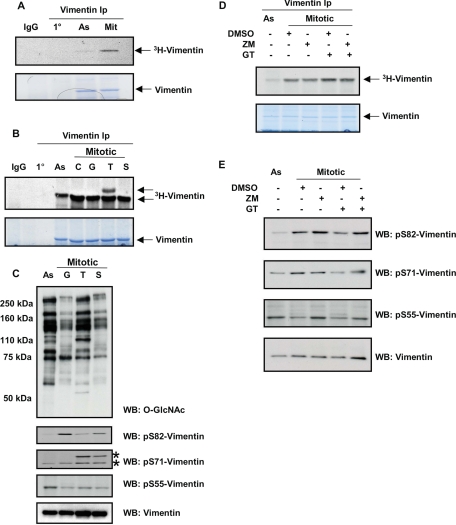
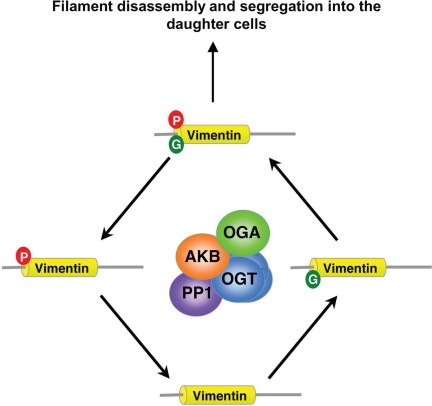
O-linked beta-N-acetylglucosamine (O-GlcNAc) is a highly dynamic intracellular protein modification responsive to stress, hormones, nutrients, and cell cycle stage. Alterations in O-GlcNAc addition or removal (cycling) impair cell cycle progression and cytokinesis, but the mechanisms are not well understood. Here, we demonstrate that the enzymes responsible for O-GlcNAc cycling, O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA) are in a transient complex at M phase with the mitotic kinase Aurora B and protein phosphatase 1. OGT colocalized to the midbody during telophase with Aurora B. Furthermore, these proteins coprecipitated with each other in a late mitotic extract. The complex was stable under Aurora inhibition; however, the total cellular levels of O-GlcNAc were increased and the localization of OGT was decreased at the midbody after Aurora inhibition. Vimentin, an intermediate filament protein, is an M phase substrate for both Aurora B and OGT. Overexpression of OGT or OGA led to defects in mitotic phosphorylation on multiple sites, whereas OGT overexpression increased mitotic GlcNAcylation of vimentin. OGA inhibition caused a decrease in vimentin late mitotic phosphorylation but increased GlcNAcylation. Together, these data demonstrate that the O-GlcNAc cycling enzymes associate with kinases and phosphatases at M phase to regulate the posttranslational status of vimentin.
Figures
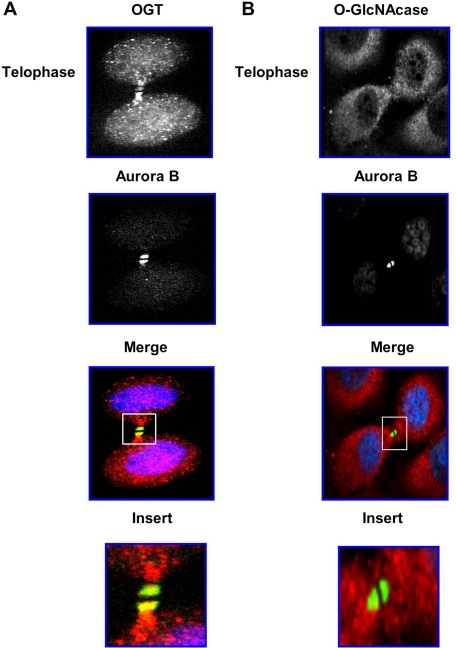
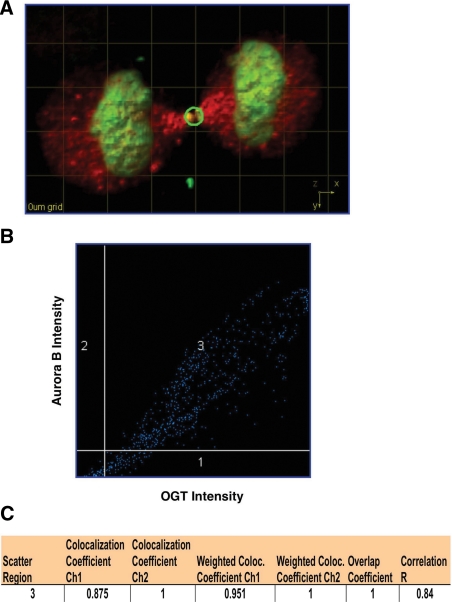
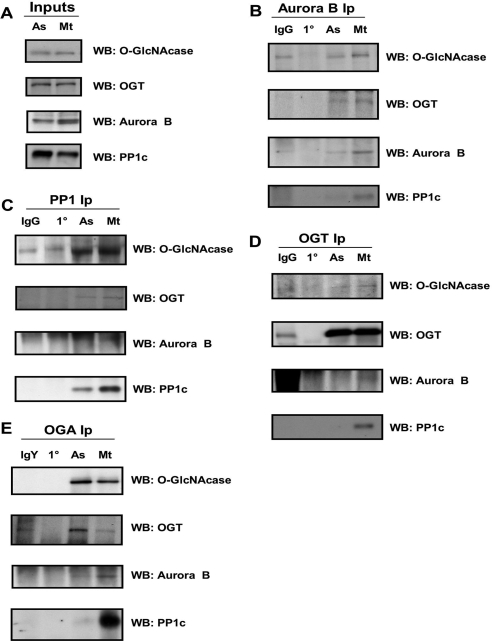
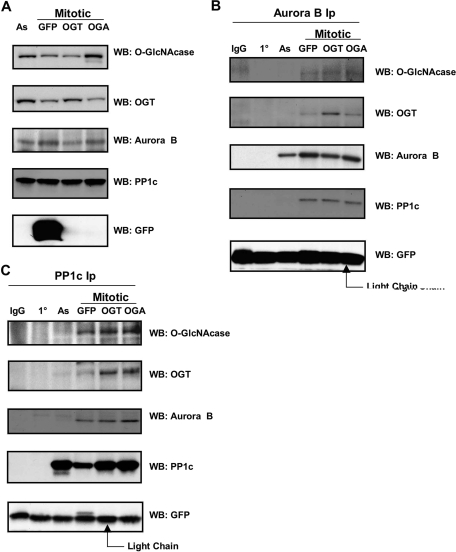
Comment in
- Mol Biol Cell. 19:4019.
References
-
- Brickley K., Smith M. J., Beck M., Stephenson F. A. GRIF-1 and OIP106, members of a novel gene family of coiled-coil domain proteins: association in vivo and in vitro with kinesin. J. Biol. Chem. 2005;280:14723–14732. - PubMed
-
- Carmena M., Earnshaw W. C. The cellular geography of aurora kinases. Nat. Rev. Mol. Cell Biol. 2003;4:842–854. - PubMed
-
- Cetinbas N., Macauley M. S., Stubbs K. A., Drapala R., Vocadlo D. J. Identification of Asp174 and Asp175 as the key catalytic residues of human O-GlcNAcase by functional analysis of site-directed mutants. Biochemistry. 2006;45:3835–3844. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
