

A positive-feedback mechanism promotes reovirus particle conversion to the intermediate associated with membrane penetration
- PMID: 18653761
- PMCID: PMC2492467
- DOI: 10.1073/pnas.0802039105
A positive-feedback mechanism promotes reovirus particle conversion to the intermediate associated with membrane penetration
Abstract
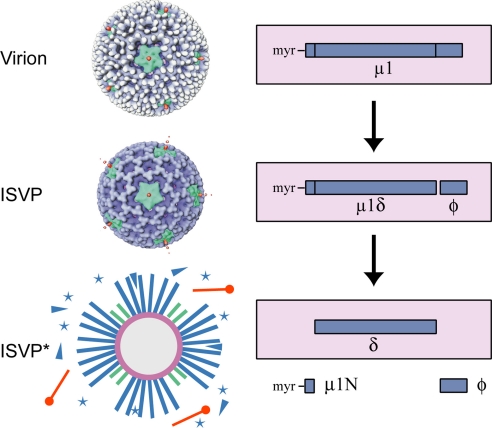
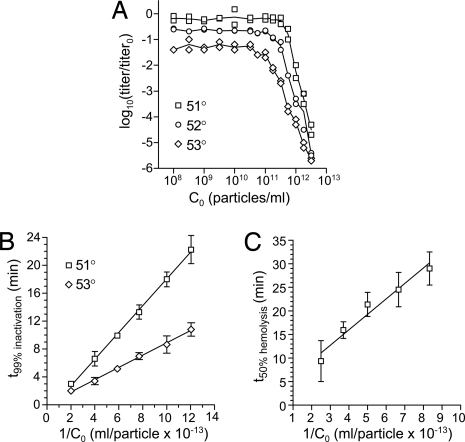
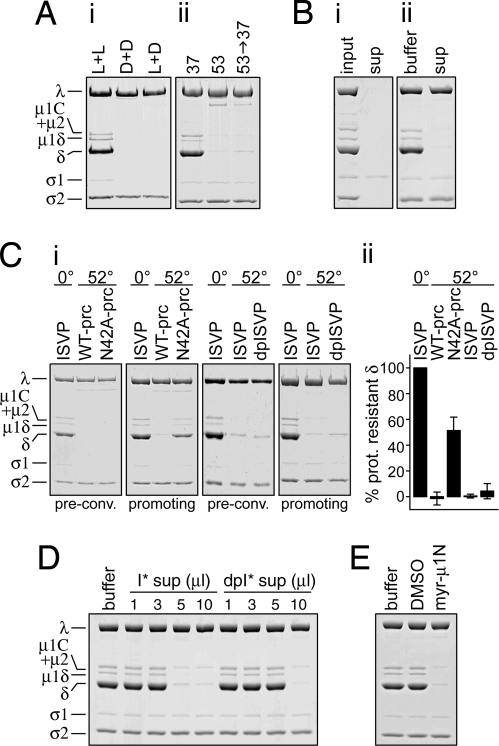
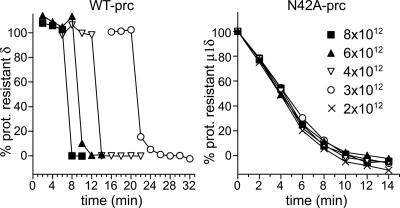
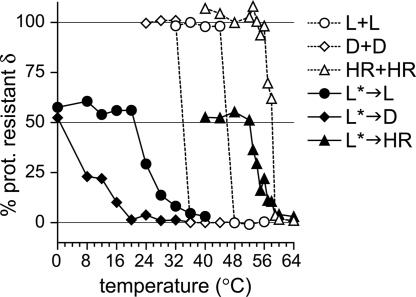
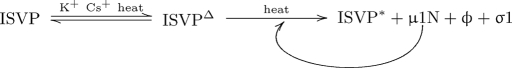
Membrane penetration by reovirus is associated with conversion of a metastable intermediate, the ISVP, to a further-disassembled particle, the ISVP*. Factors that promote this conversion in cells are poorly understood. Here, we report the in vitro characterization of a positive-feedback mechanism for promoting ISVP* conversion. At high particle concentration, conversion approximated second-order kinetics, and products of the reaction operated in trans to promote the conversion of target ISVPs. Pore-forming peptide mu1N, which is released from particles during conversion, was sufficient for promoting activity. A mutant that does not undergo mu1N release failed to exhibit second-order conversion kinetics and also failed to promote conversion of wild-type target ISVPs. Susceptibility of target ISVPs to promotion in trans was temperature dependent and correlated with target stability, suggesting that capsid dynamics are required to expose the interacting epitope. A positive-feedback mechanism of promoting escape from the metastable intermediate has not been reported for other viruses but represents a generalizable device for sensing a confined volume, such as that encountered during cell entry.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
