

Phosphate and pyrophosphate mediate PKA-induced vascular cell calcification
- PMID: 18655772
- PMCID: PMC2570192
- DOI: 10.1016/j.bbrc.2008.07.062
Phosphate and pyrophosphate mediate PKA-induced vascular cell calcification
Abstract
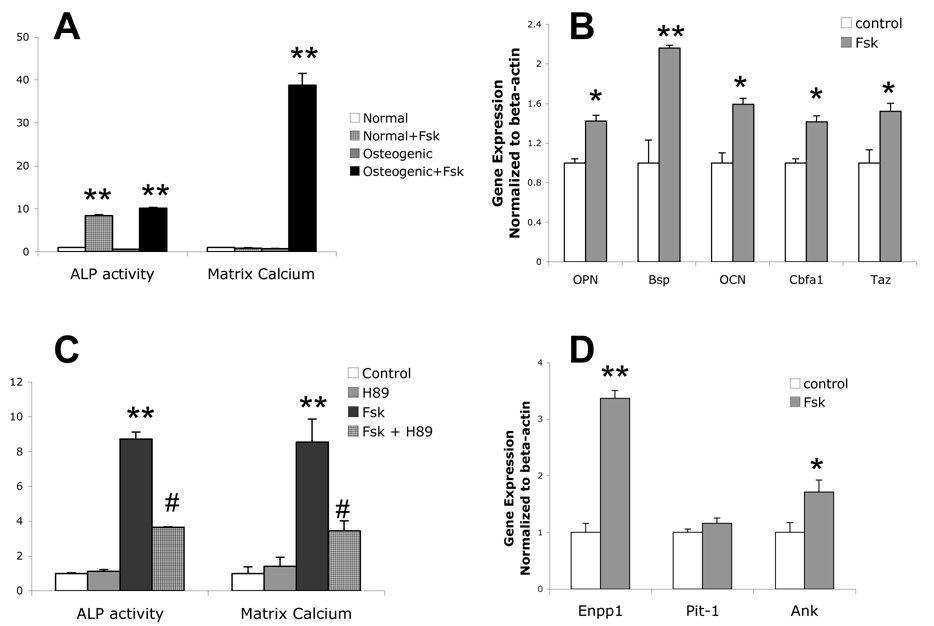
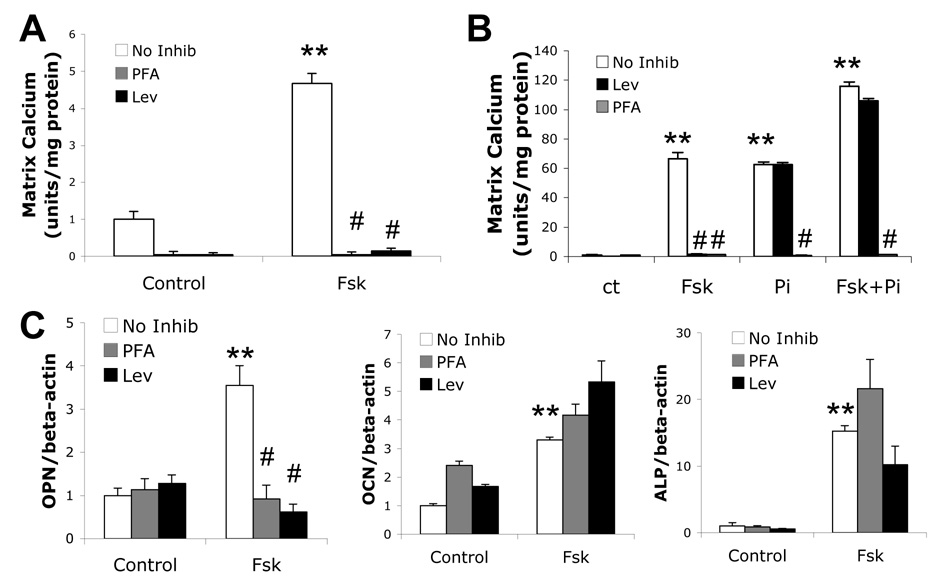
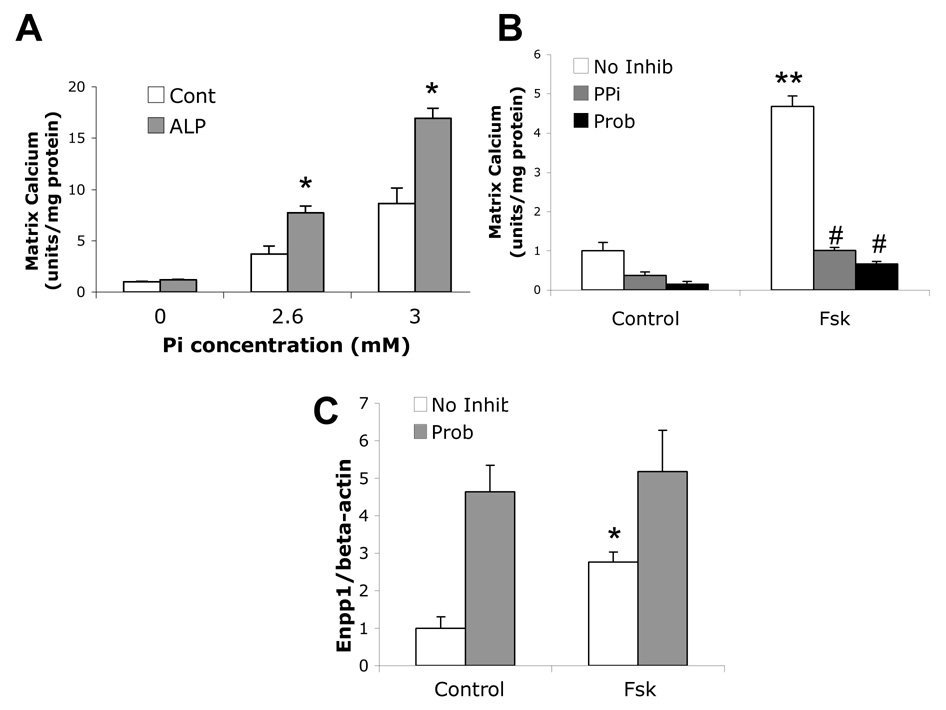
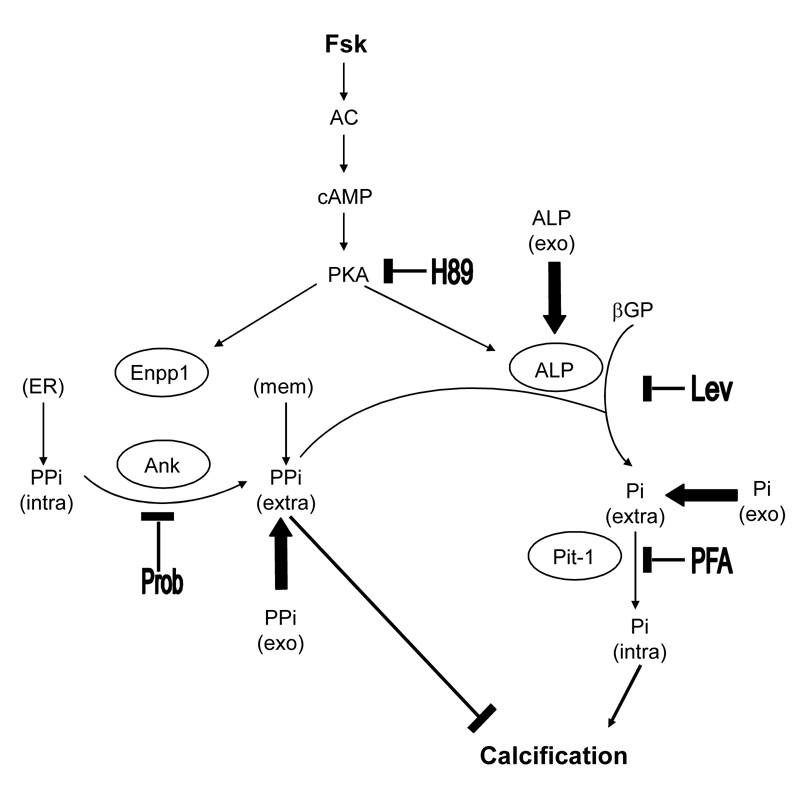
Vascular calcification is associated with increased cardiovascular risk and occurs by osteochondrogenic differentiation of vascular cells. Many of the same regulatory factors that control skeletal mineralization, including the complex metabolic pathway controlling levels of the activator, inorganic phosphate, and the potent inhibitor, pyrophosphate, also govern vascular calcification. We previously found that the cAMP/PKA signaling pathway mediates in vitro vascular cell calcification induced by inflammatory factors including tumor necrosis factor-alpha 1 and oxidized phospholipids. In this report, we tested whether this signaling pathway modulates phosphate and pyrophosphate metabolism. Treatment of primary murine aortic cells with the PKA activator, forskolin, significantly induced osteoblastic differentiation markers, including alkaline phosphatase (ALP), osteopontin, and osteocalcin as well as the pyrophosphate generator, ectonucleotide-pyrophosphatase/phosphodiesterase-1 (Enpp1) and the pyrophosphate transporter, ankylosis protein, but not the sodium/phosphate cotransporter, Pit-1. In the presence of a substrate for ALP, beta-glycerophosphate, which generates inorganic phosphate, forskolin also enhanced matrix mineralization. Inhibitors of ALP or Pit-1 abrogated forskolin-induced osteopontin expression and mineralization but not forskolin-induced osteocalcin or ALP. These results suggest that phosphate is necessary for PKA-induced calcification of vascular cells and that the extent of PKA-induced calcification is controlled by feedback induction of the inhibitor, pyrophosphate.
Figures
References
-
- Jeziorska M, Woolley DE. Neovascularization in early atherosclerotic lesions of human carotid arteries: its potential contribution to plaque development. Hum Pathol. 1999;30:919–925. - PubMed
-
- Jeziorska M, Woolley DE. Local neovascularization and cellular composition within vulnerable regions of atherosclerotic plaques of human carotid arteries. J Pathol. 1999;188:189–196. - PubMed
-
- Stary HC. Natural history of calcium deposits in atherosclerosis progression and regression. Z Kardiol. 2000;89 Suppl 2:28–35. - PubMed
-
- Abedin M, Lim J, Tang TB, Park D, Demer LL, Tintut Y. N-3 fatty acids inhibit vascular calcification via the p38-mitogen-activated protein kinase and peroxisome proliferator-activated receptor-gamma pathways. Circ Res. 2006;98:727–729. - PubMed
-
- Moe SM, Chen NX. Pathophysiology of vascular calcification in chronic kidney disease. Circ Res. 2004;95:560–567. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
