

Potential utility of aquaporin modulators for therapy of brain disorders
- PMID: 18655912
- PMCID: PMC3601944
- DOI: 10.1016/S0079-6123(08)00446-9
Potential utility of aquaporin modulators for therapy of brain disorders
Abstract
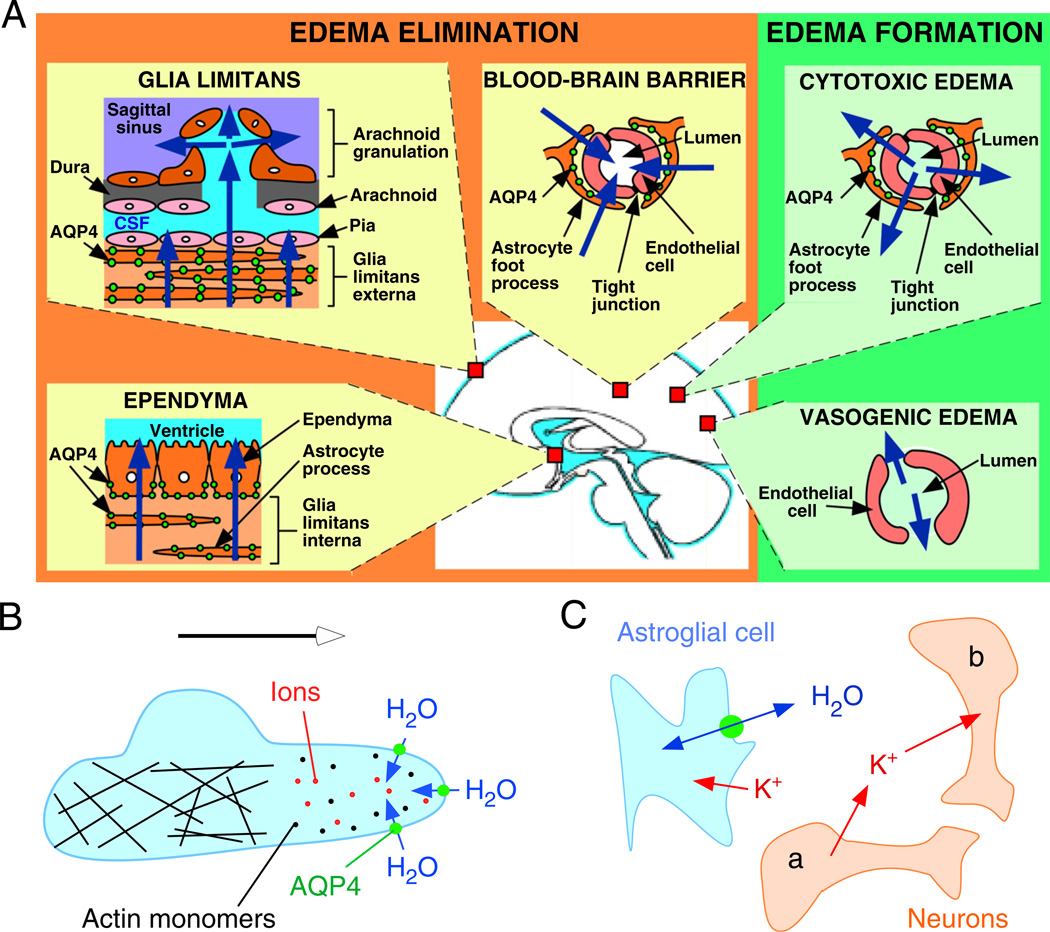
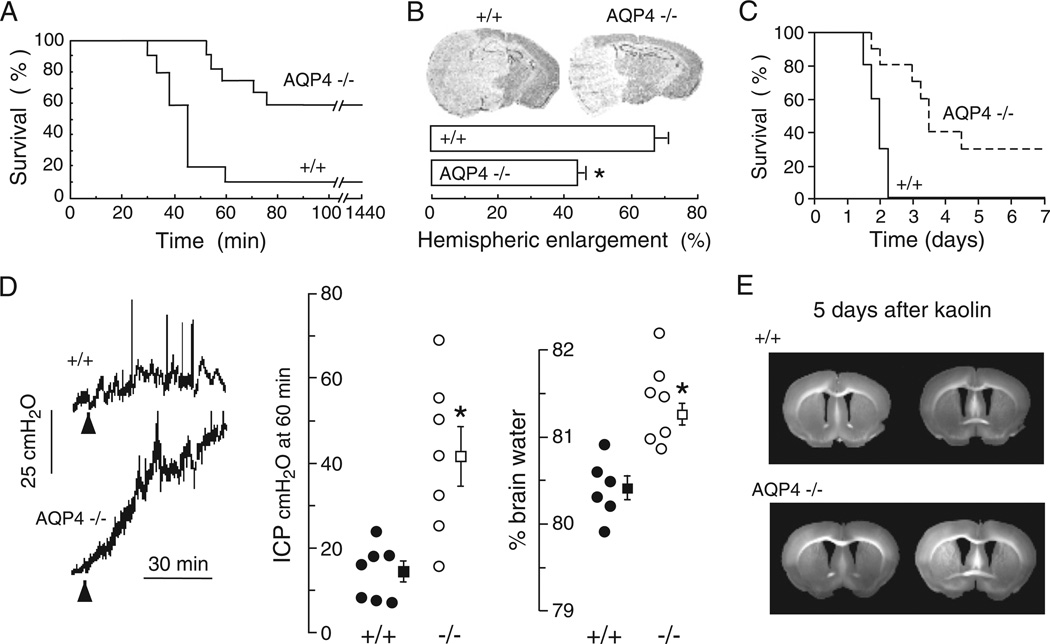
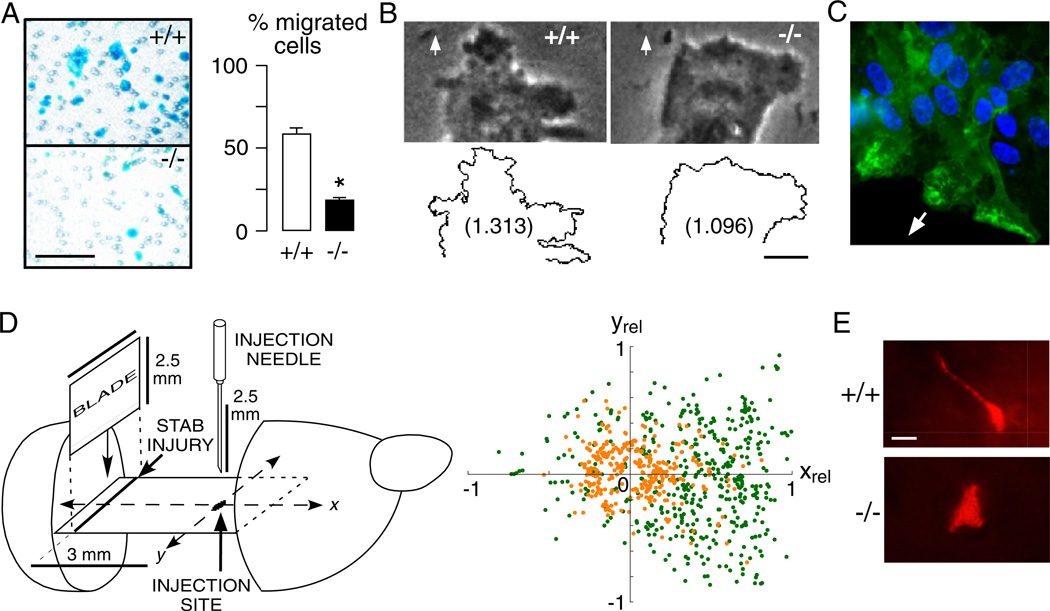
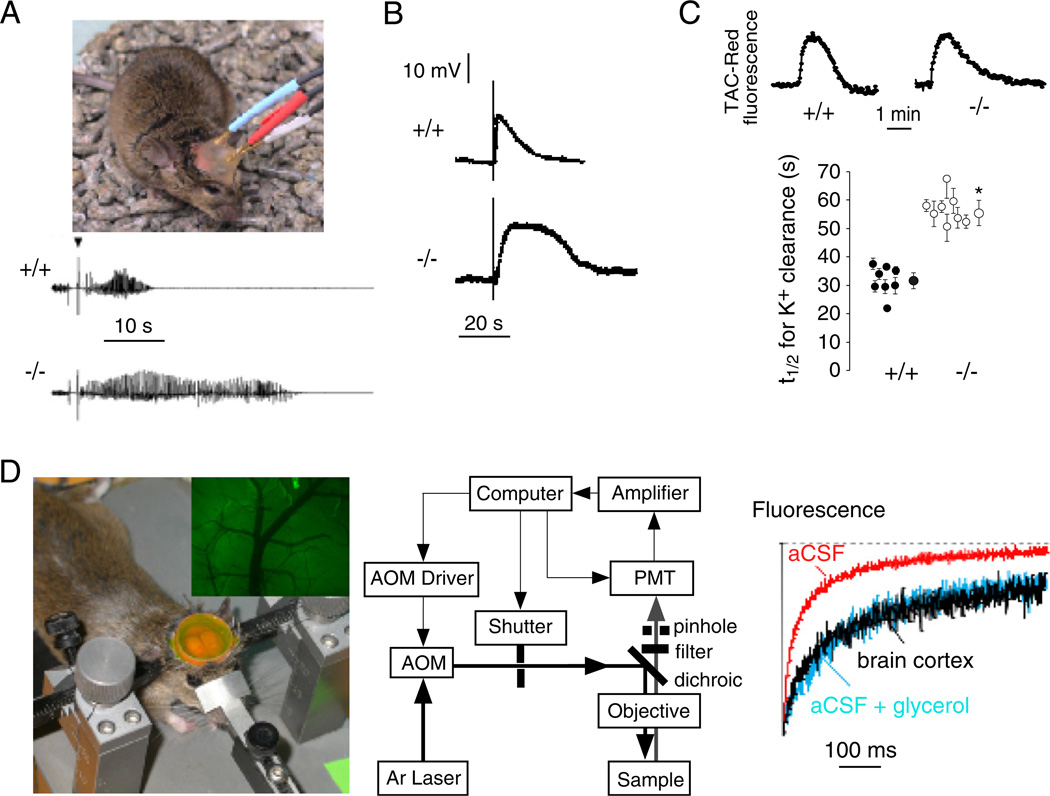
Of the several aquaporin (AQP) water channels expressed in the central nervous system, AQP4 is an attractive target for drug discovery. AQP4 is expressed in astroglia, most strongly at the blood-brain and brain-cerebrospinal fluid barriers. Phenotype analysis of AQP4 knockout mice indicates the involvement of AQP4 in three distinct processes: brain water balance, astroglial cell migration and neural signal transduction. By slowing water uptake into the brain, AQP4 knockout mice manifest reduced brain swelling and improved outcome in models of cytotoxic cerebral oedema such as water intoxication, focal ischaemia and meningitis. However, by slowing the clearance of excess water from brain, AQP4 knockout mice do worse in models of vasogenic oedema such as brain tumour, abscess and hydrocephalus. AQP4 deficient astroglial cells show greatly impaired migration in response to chemotactic stimuli, reducing glial scar formation, by a mechanism that we propose involves AQP4-facilitated water flux in lamellipodia of migrating cells. AQP4 knockout mice also manifest increased seizure threshold and duration, by a mechanism that may involve slowed K(+) uptake from the extracellular space (ECS) following neuroexcitation, as well as ECS expansion. Notwithstanding challenges in drug delivery to the central nervous system and their multiplicity of actions, AQP4 inhibitors have potential utility in reducing cytotoxic brain swelling, increasing seizure threshold and reducing glial scar formation; enhancers of AQP4 expression have potential utility in reducing vasogenic brain swelling. AQP4 modulators may thus offer new therapeutic options for stroke, tumour, infection, hydrocephalus, epilepsy and traumatic brain and spinal cord injury.
Figures
References
-
- Amiry-Moghaddam M, Lindland H, Zelenin S, Roberg BA, Gundersen BB, Petersen P, Rinvik E, Torgner IA, Ottersen OP. Brain mitochondria contain aquaporin water channels: evidence for the expression of a short AQP9 isoform in the inner mitochondrial membrane. Faseb J. 2005;19:1459–1467. - PubMed
-
- Amiry-Moghaddam M, Otsuka T, Hurn PD, Traystman RJ, Haug FM, Froehner SC, Adams ME, Neely JD, Agre P, Ottersen OP, Bhardwaj A. An alpha-syntrophin-dependent pool of AQP4 in astroglial end-feet confers bidirectional water flow between blood and brain. Proc. Natl. Acad. Sci. U.S.A. 2003a;100:2106–2111. - PMC - PubMed
-
- Amiry-Moghaddam M, Williamson A, Palomba M, Eid T, de Lanerolle NC, Nagelhus EA, Adams ME, Froehner SC, Agre P, Ottersen OP. Delayed K+ clearance associated with aquaporin-4 mislocalization: phenotypic defects in brains of alpha-syntrophin-null mice. Proc. Natl. Acad. Sci. U.S.A. 2003b;100:13615–13620. - PMC - PubMed
-
- Amiry-Moghaddam M, Xue R, Haug FM, Neely JD, Bhardwaj A, Agre P, Adams ME, Froehner SC, Mori S, Ottersen OP. Alpha-syntrophin deletion removes the perivascular but not endothelial pool of aquaporin-4 at the blood–brain barrier and delays the development of brain edema in an experimental model of acute hyponatremia. Faseb J. 2004;18:542–544. - PubMed
-
- Aoki K, Uchihara T, Tsuchiya K, Nakamura A, Ikeda K, Wakayama Y. Enhanced expression of aquaporin 4 in human brain with infarction. Acta Neuropathol. (Berl.) 2003;106:121–124. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
