

From fish to modern humans--comparative anatomy, homologies and evolution of the head and neck musculature
- PMID: 18657257
- PMCID: PMC2644766
- DOI: 10.1111/j.1469-7580.2008.00953.x
From fish to modern humans--comparative anatomy, homologies and evolution of the head and neck musculature
Abstract
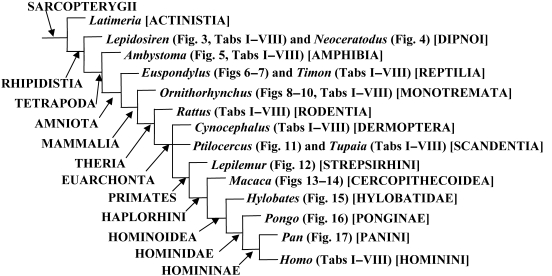
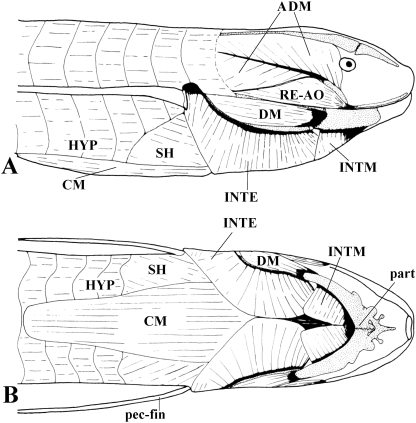
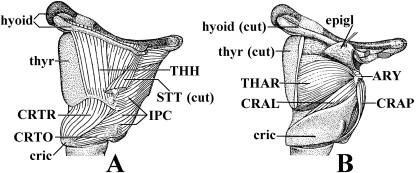
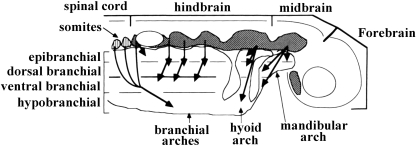
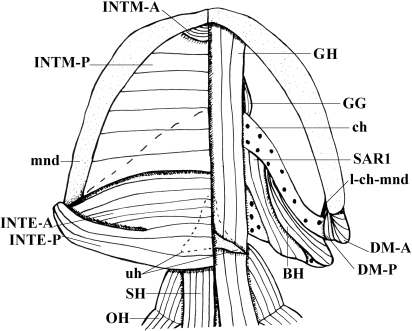
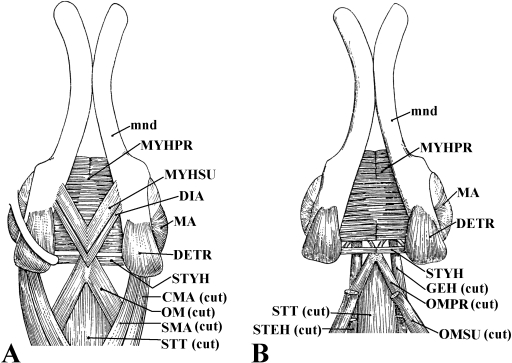
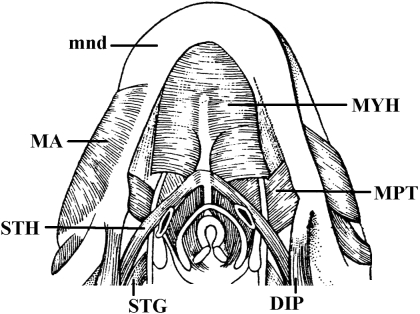
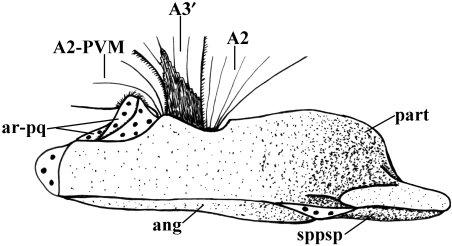
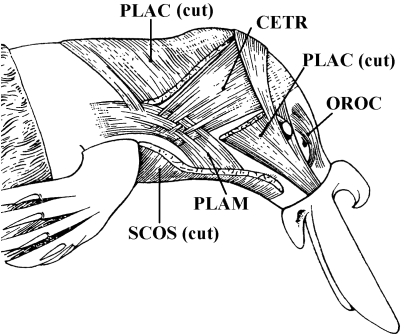
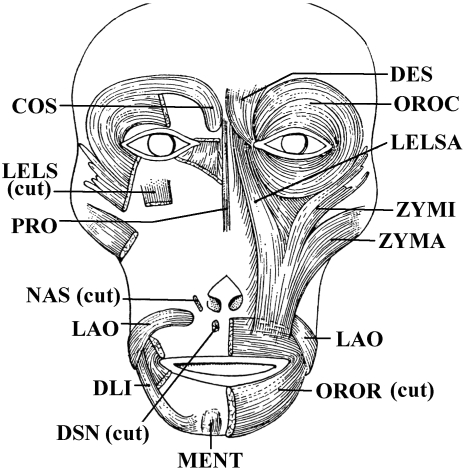
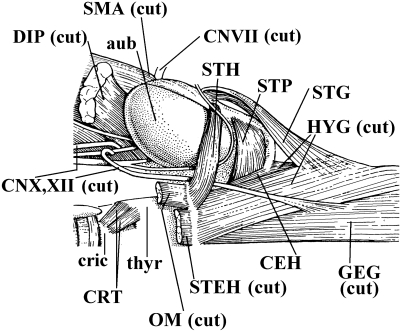
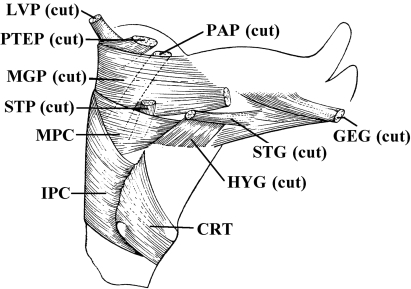
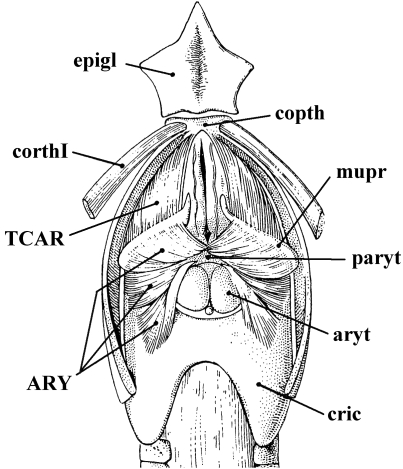
In a recent paper Diogo (2008) reported the results of the first part of an investigation of the comparative anatomy, homologies and evolution of the head and neck muscles of osteichthyans (bony fish + tetrapods). That report mainly focused on actinopterygian fish, but also compared these fish with certain non-mammalian sarcopterygians. The present paper focuses mainly on sarcopterygians, and particularly on how the head and neck muscles have evolved during the transitions from sarcopterygian fish and non-mammalian tetrapods to monotreme and therian mammals, including modern humans. The data obtained from our dissections of the head and neck muscles of representative members of sarcopterygian fish, amphibians, reptiles, monotremes and therian mammals, such as rodents, tree-shrews, colugos and primates, including modern humans, are compared with the information available in the literature. Our observations and comparisons indicate that the number of mandibular and true branchial muscles (sensu this work) present in modern humans is smaller than that found in mammals such as tree-shrews, rats and monotremes, as well as in reptiles such as lizards. Regarding the pharyngeal musculature, there is an increase in the number of muscles at the time of the evolutionary transition leading to therian mammals, but there was no significant increase during the transition leading to the emergence of higher primates and modern humans. The number of hypobranchial muscles is relatively constant within the therian mammals we examined, although in this case modern humans have more muscles than other mammals. The number of laryngeal and facial muscles in modern humans is greater than that found in most other therian taxa. Interestingly, modern humans possess peculiar laryngeal and facial muscles that are not present in the majority of the other mammalian taxa; this seems to corroborate the crucial role played by vocal communication and by facial expressions in primate and especially in human evolution. It is hoped that by compiling, in one paper, data about the head and neck muscles of a wide range of sarcopterygians, the present work could be useful to comparative anatomists, evolutionary biologists and functional morphologists and to researchers working in other fields such as developmental biology, genetics and/or evolutionary developmental biology.
Figures
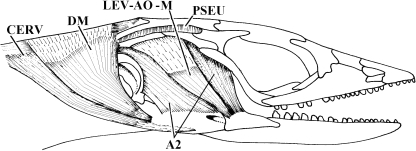
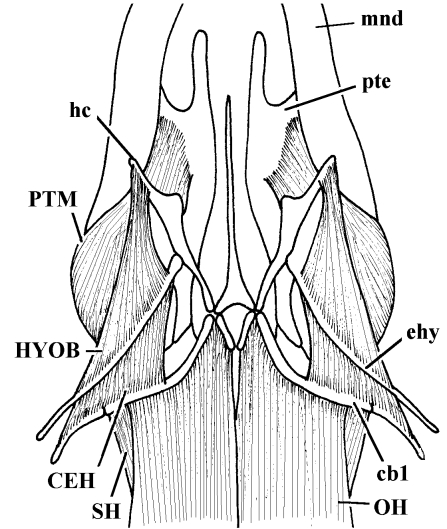
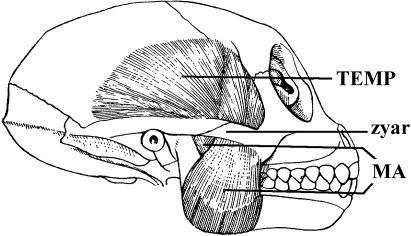
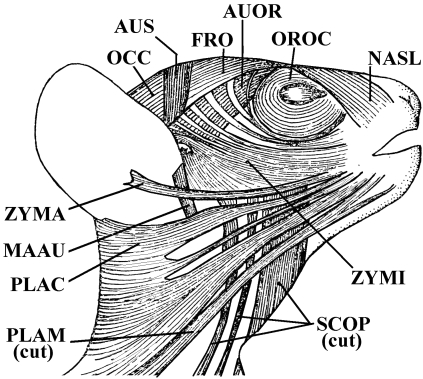
Similar articles
-
Soft-tissue anatomy of the primates: phylogenetic analyses based on the muscles of the head, neck, pectoral region and upper limb, with notes on the evolution of these muscles.J Anat. 2011 Sep;219(3):273-359. doi: 10.1111/j.1469-7580.2011.01403.x. Epub 2011 Jun 20. J Anat. 2011. PMID: 21689100 Free PMC article.
-
From fish to modern humans--comparative anatomy, homologies and evolution of the pectoral and forelimb musculature.J Anat. 2009 May;214(5):694-716. doi: 10.1111/j.1469-7580.2009.01067.x. J Anat. 2009. PMID: 19438764 Free PMC article.
-
Development of mandibular, hyoid and hypobranchial muscles in the zebrafish: homologies and evolution of these muscles within bony fishes and tetrapods.BMC Dev Biol. 2008 Feb 28;8:24. doi: 10.1186/1471-213X-8-24. BMC Dev Biol. 2008. PMID: 18307809 Free PMC article.
-
On the origin, homologies and evolution of primate facial muscles, with a particular focus on hominoids and a suggested unifying nomenclature for the facial muscles of the Mammalia.J Anat. 2009 Sep;215(3):300-19. doi: 10.1111/j.1469-7580.2009.01111.x. Epub 2009 Jun 12. J Anat. 2009. PMID: 19531159 Free PMC article. Review.
-
Specialize or risk disappearance - empirical evidence of anisomerism based on comparative and developmental studies of gnathostome head and limb musculature.Biol Rev Camb Philos Soc. 2015 Aug;90(3):964-78. doi: 10.1111/brv.12142. Epub 2014 Aug 30. Biol Rev Camb Philos Soc. 2015. PMID: 25174804 Review.
Cited by
-
Raising an Eye at Facial Muscle Morphology in Canids.Biology (Basel). 2024 Apr 25;13(5):290. doi: 10.3390/biology13050290. Biology (Basel). 2024. PMID: 38785773 Free PMC article.
-
Facial display and blushing: Means of visual communication in blue-and-yellow macaws (Ara Ararauna)?PLoS One. 2018 Aug 22;13(8):e0201762. doi: 10.1371/journal.pone.0201762. eCollection 2018. PLoS One. 2018. PMID: 30133471 Free PMC article.
-
Soft-tissue anatomy of the primates: phylogenetic analyses based on the muscles of the head, neck, pectoral region and upper limb, with notes on the evolution of these muscles.J Anat. 2011 Sep;219(3):273-359. doi: 10.1111/j.1469-7580.2011.01403.x. Epub 2011 Jun 20. J Anat. 2011. PMID: 21689100 Free PMC article.
-
Dog facial landmarks detection and its applications for facial analysis.Sci Rep. 2025 Jul 1;15(1):21886. doi: 10.1038/s41598-025-07040-3. Sci Rep. 2025. PMID: 40595051 Free PMC article.
-
Convergent evolution of a mobile bony tongue in flighted dinosaurs and pterosaurs.PLoS One. 2018 Jun 20;13(6):e0198078. doi: 10.1371/journal.pone.0198078. eCollection 2018. PLoS One. 2018. PMID: 29924798 Free PMC article.
References
-
- Abdala V, Moro S. A cladistic analysis of ten lizard families (Reptilia: Squamata) based on cranial musculature. Russ J Herpetol. 2003;10:53–78.
-
- Adamicka P, Ahnelt H. Two jaw articulations in Latimeria chalumnae(Actinistia, Coelacanthidae) Zool Jb Anat. 1992;122:107–112.
-
- Alexander RMcN. Jaw mechanisms of the coelacanth Latimeria. Copeia. 1973;1973:156–158.
-
- Allis EP. The cranial muscles and cranial nerves of Amia calva. J Morphol. 1897;12:487–737.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
