

Structure of the myristylated human immunodeficiency virus type 2 matrix protein and the role of phosphatidylinositol-(4,5)-bisphosphate in membrane targeting
- PMID: 18657545
- PMCID: PMC2581411
- DOI: 10.1016/j.jmb.2008.07.027
Structure of the myristylated human immunodeficiency virus type 2 matrix protein and the role of phosphatidylinositol-(4,5)-bisphosphate in membrane targeting
Abstract
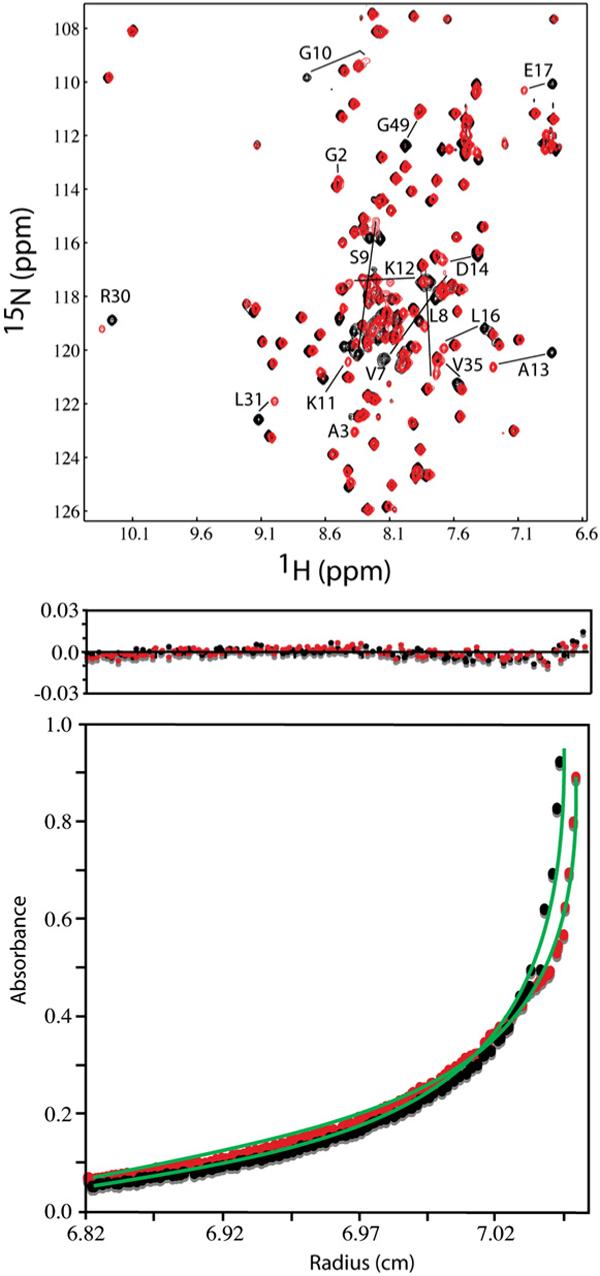
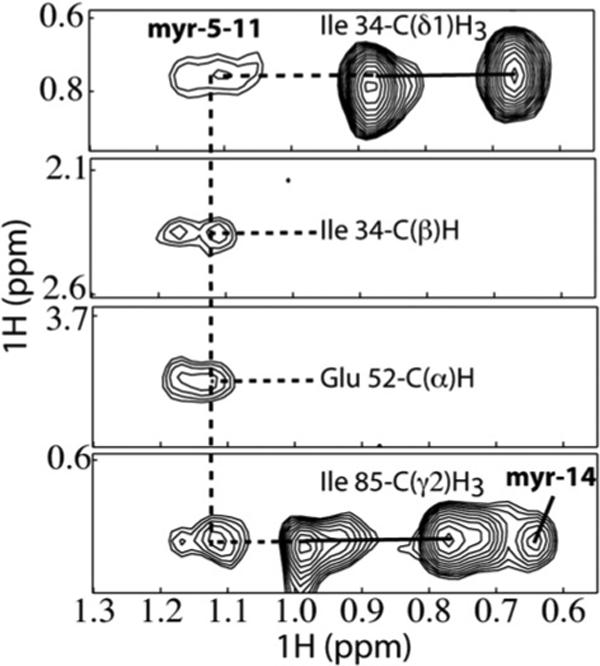
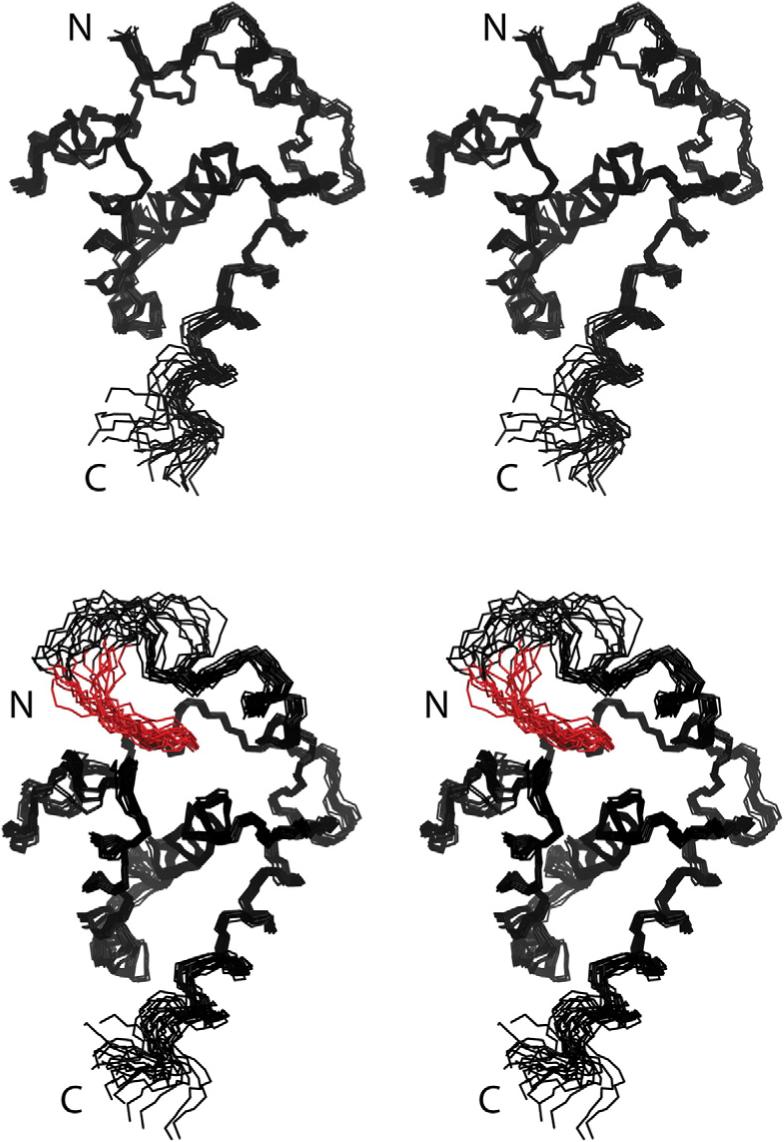
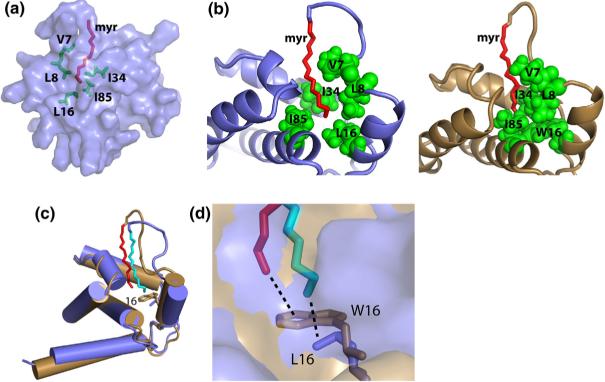
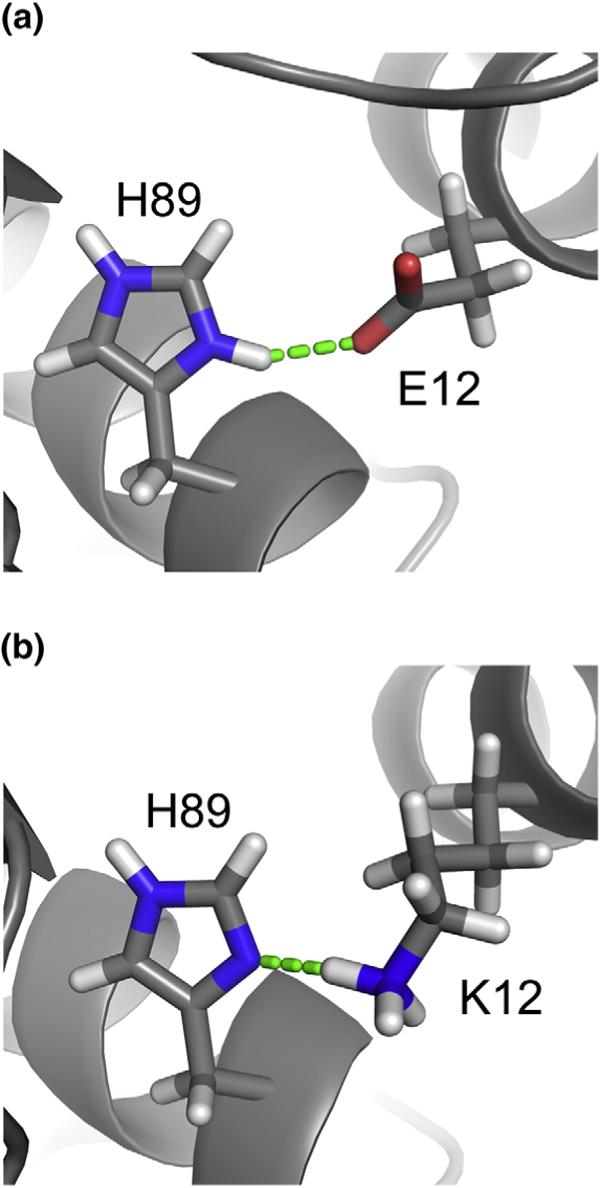
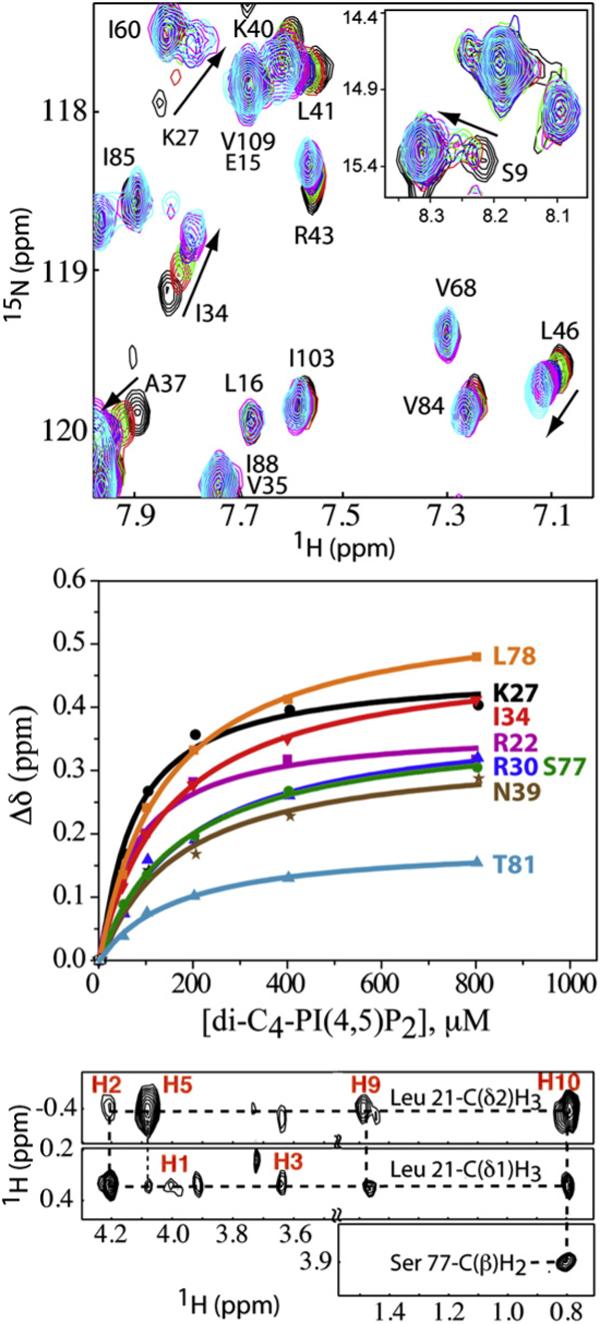
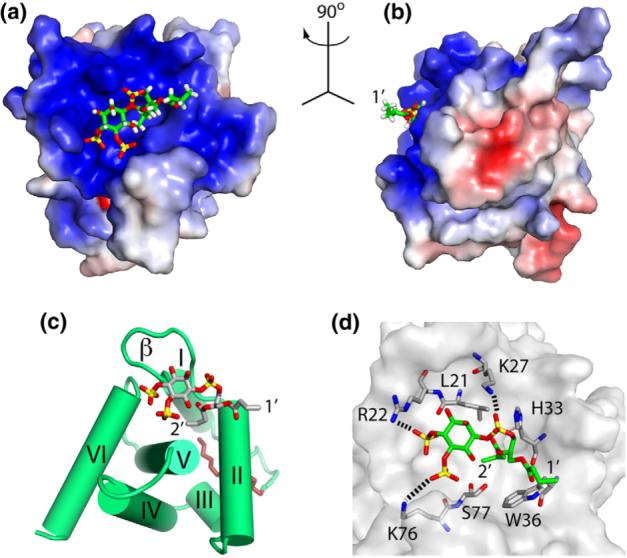
During the late phase of retroviral replication, newly synthesized Gag proteins are targeted to the plasma membrane (PM), where they assemble and bud to form immature virus particles. Membrane targeting by human immunodeficiency virus type 1 (HIV-1) Gag is mediated by the PM marker molecule phosphatidylinositol-(4,5)-bisphosphate [PI(4,5)P(2)], which is capable of binding to the matrix (MA) domain of Gag in an extended lipid conformation and of triggering myristate exposure. Here, we show that, as observed previously for HIV-1 MA, the myristyl group of HIV-2 MA is partially sequestered within a narrow hydrophobic tunnel formed by side chains of helices 1, 2, 3, and 5. However, the myristate of HIV-2 MA is more tightly sequestered than that of the HIV-1 protein and does not exhibit concentration-dependent exposure. Soluble PI(4,5)P(2) analogs containing truncated acyl chains bind HIV-2 MA and induce minor long-range structural changes but do not trigger myristate exposure. Despite these differences, the site of HIV-2 assembly in vivo can be manipulated by enzymes that regulate PI(4,5)P(2) localization. Our findings indicate that HIV-1 and HIV-2 are both targeted to the PM for assembly via a PI(4,5)P(2)-dependent mechanism, despite differences in the sensitivity of the MA myristyl switch, and suggest a potential mechanism that may contribute to the poor replication kinetics of HIV-2.
Figures


References
-
- Poulsen AG, Aaby P, Larsen O, Jensen H, Naucler A, Lisse IM, et al. 9-year HIV-2-associated mortality in an urban community in Bissau, West Africa. Lancet. 1997;349:911–914. - PubMed
-
- Schim van der Loeff MF, Jaffar S, Aveika AA, Sabally S, Corrah T, Harding E, et al. Mortality of HIV-1, HIV-2 and HIV-1/HIV-2 dually infected patients in a clinic-based cohort in The Gambia. AIDS. 2002;16:1775–1783. - PubMed
-
- Ariyoshi K, Jaffar S, Alabi AS, Berry N, Schim van der Loeff MF, Sabally S, et al. Plasma RNA viral load predicts the rate of CD4 T cell decline and death in HIV-2-infected patients in West Africa. AIDS. 2000;14:339–344. - PubMed
-
- Mellors JW, Rinaldo CR, Gupta P, Jr, White RM, Todd JA, Kingsley LA. Prognosis in HIV-1 infection predicted by the quantity of virus in plasma. Science. 1996;272:1167–1170. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
