

Experimental verification of the behavioral foundation of bacterial transport parameters using microfluidics
- PMID: 18658218
- PMCID: PMC2567943
- DOI: 10.1529/biophysj.108.134510
Experimental verification of the behavioral foundation of bacterial transport parameters using microfluidics
Abstract
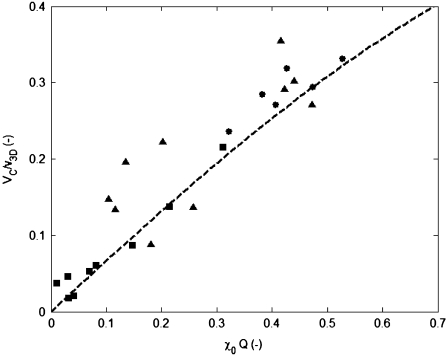
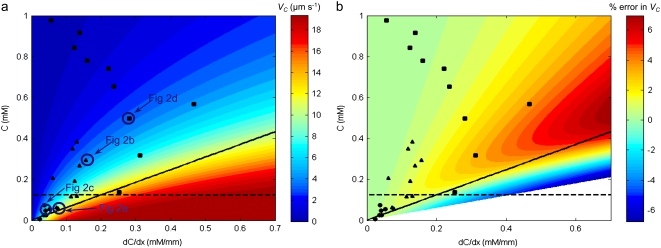
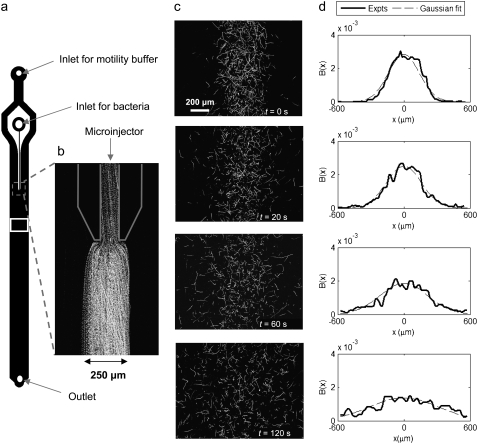
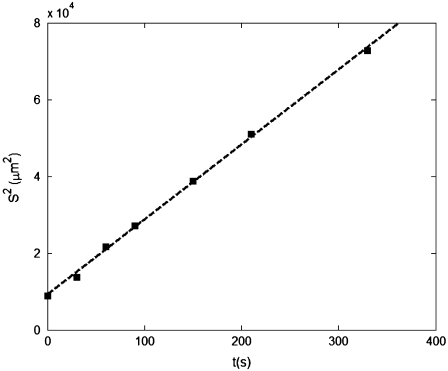
We present novel microfluidic experiments to quantify population-scale transport parameters (chemotactic sensitivity chi(0) and random motility mu) of a population of bacteria. Previously, transport parameters have been derived theoretically from single-cell swimming behavior using probabilistic models, yet the mechanistic foundations of this upscaling process have not been verified experimentally. We designed a microfluidic capillary assay to generate and accurately measure gradients of chemoattractant (alpha-methylaspartate) while simultaneously capturing the swimming trajectories of individual Escherichia coli bacteria using videomicroscopy and cell tracking. By measuring swimming speed and bias in the swimming direction of single cells for a range of chemoattractant concentrations and concentration gradients, we directly computed the chemotactic velocity VC and the associated chemotactic sensitivity chi(0). We then show how mu can also be readily determined using microfluidics but that a population-scale microfluidic approach is experimentally more convenient than a single-cell analysis in this case. Measured values of both chi(0) [(12.4 +/- 2.0) x 10(-4) cm(2) s(-1)] and mu [(3.3 +/- 0.8) x 10(-6) cm(2) s(-1)] are comparable to literature results. This microscale approach to bacterial chemotaxis lends experimental support to theoretical derivations of population-scale transport parameters from single-cell behavior. Furthermore, this study shows that microfluidic platforms can go beyond traditional chemotaxis assays and enable the quantification of bacterial transport parameters.
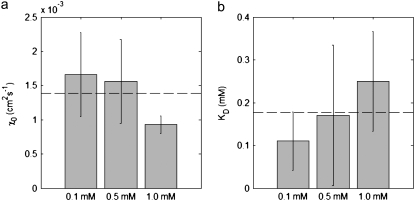
Figures
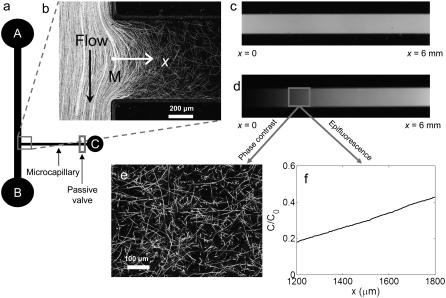
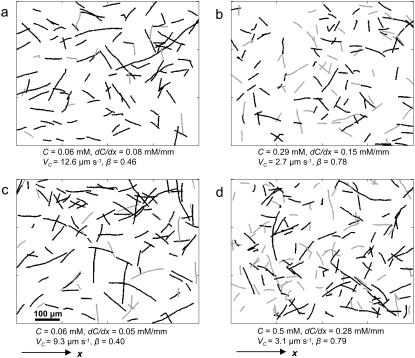
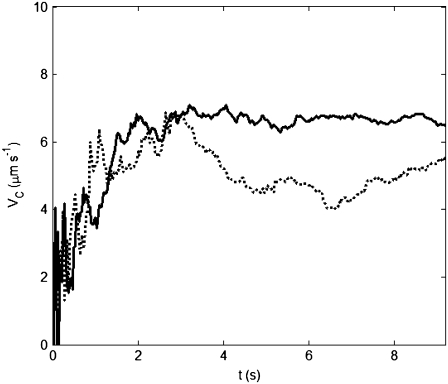
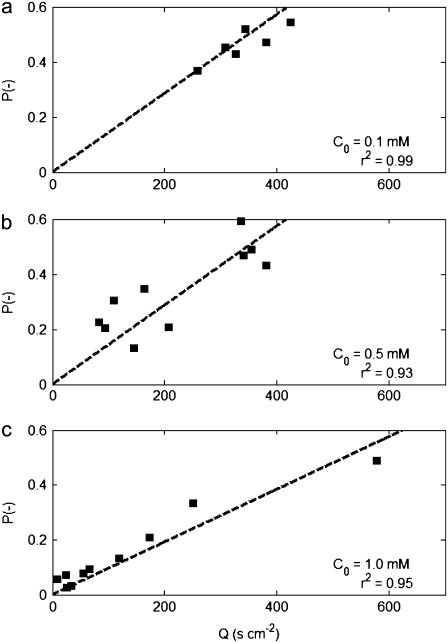
References
-
- Koyama, S., D. Amarie, H. A. Soini, M. V. Novotny, and S. C. Jacobson. 2006. Chemotaxis assays of mouse sperm on microfluidic devices. Anal. Chem. 78:3354–3359. - PubMed
-
- Fauci, L. J., and R. Dillon. 2006. Biofluidmechanics of reproduction. Annu. Rev. Fluid Mech. 38:371–394.
-
- Pratt, L. A., and R. Kolter. 1998. Genetic analysis of Escherichia coli biofilm formation: roles of flagella, motility, chemotaxis and type I pili. Mol. Microbiol. 30:285–293. - PubMed
-
- O'Toole, G., H. B. Kaplan, and R. Kolter. 2000. Biofilm formation as microbial development. Annu. Rev. Microbiol. 54:49–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
