

Heterotrimeric G-protein subunit function in Candida albicans: both the alpha and beta subunits of the pheromone response G protein are required for mating
- PMID: 18658257
- PMCID: PMC2547078
- DOI: 10.1128/EC.00077-08
Heterotrimeric G-protein subunit function in Candida albicans: both the alpha and beta subunits of the pheromone response G protein are required for mating
Abstract
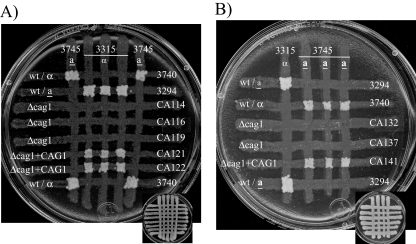
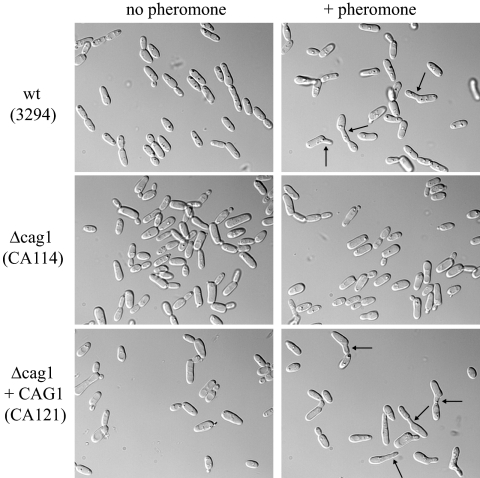
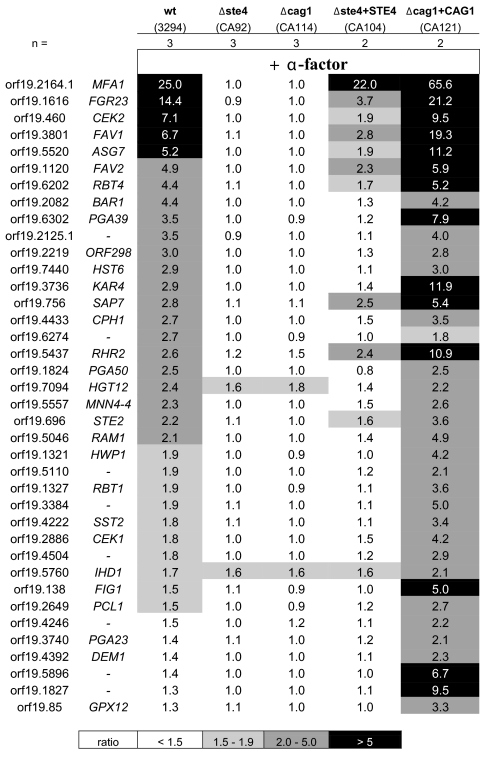
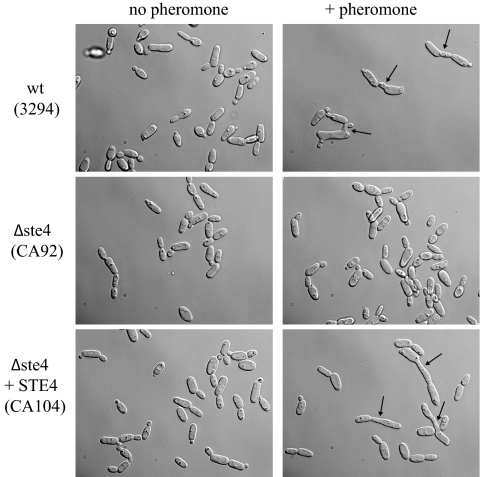
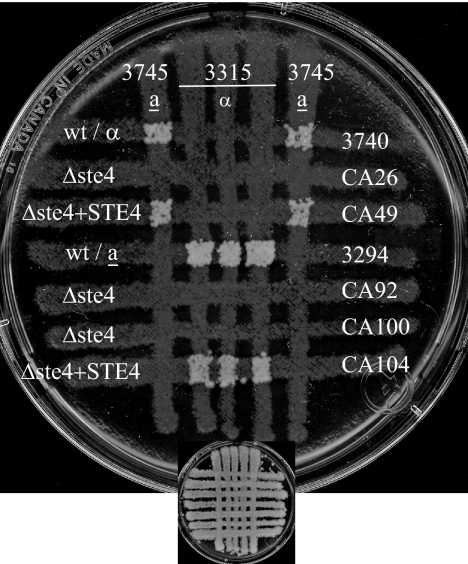
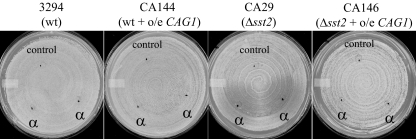
A pheromone-mediated signaling pathway that couples seven-transmembrane-domain (7-TMD) receptors to a mitogen-activated protein kinase module controls Candida albicans mating. 7-TMD receptors are typically connected to heterotrimeric G proteins whose activation regulates downstream effectors. Two Galpha subunits in C. albicans have been identified previously, both of which have been implicated in aspects of pheromone response. Cag1p was found to complement the mating pathway function of the pheromone receptor-coupled Galpha subunit in Saccharomyces cerevisiae, and Gpa2p was shown to have a role in the regulation of cyclic AMP signaling in C. albicans and to repress pheromone-mediated arrest. Here, we show that the disruption of CAG1 prevented mating, inactivated pheromone-mediated arrest and morphological changes, and blocked pheromone-mediated gene expression changes in opaque cells of C. albicans and that the overproduction of CAG1 suppressed the hyperactive cell cycle arrest exhibited by sst2 mutant cells. Because the disruption of the STE4 homolog constituting the only C. albicans gene for a heterotrimeric Gbeta subunit also blocked mating and pheromone response, it appears that in this fungal pathogen the Galpha and Gbeta subunits do not act antagonistically but, instead, are both required for the transmission of the mating signal.
Figures
Similar articles
-
The Gbeta(KlSte4p) subunit of the heterotrimeric G protein has a positive and essential role in the induction of mating in the yeast Kluyveromyces lactis.Yeast. 2005 Sep;22(12):947-56. doi: 10.1002/yea.1278. Yeast. 2005. PMID: 16134098
-
A Candida albicans-specific region of the alpha-pheromone receptor plays a selective role in the white cell pheromone response.Mol Microbiol. 2009 Feb;71(4):925-47. doi: 10.1111/j.1365-2958.2008.06575.x. Epub 2008 Dec 18. Mol Microbiol. 2009. PMID: 19170873
-
Receptor inhibition of pheromone signaling is mediated by the Ste4p Gbeta subunit.Mol Cell Biol. 1999 Jan;19(1):441-9. doi: 10.1128/MCB.19.1.441. Mol Cell Biol. 1999. PMID: 9858568 Free PMC article.
-
Mating in Candida albicans and the search for a sexual cycle.Annu Rev Microbiol. 2005;59:233-55. doi: 10.1146/annurev.micro.59.030804.121310. Annu Rev Microbiol. 2005. PMID: 15910278 Review.
-
Analogous telesensing pathways regulate mating and virulence in two opportunistic human pathogens.mBio. 2010 Sep 7;1(4):e00181-10. doi: 10.1128/mBio.00181-10. mBio. 2010. PMID: 20827374 Free PMC article. Review.
Cited by
-
Genome Wide Analysis of WD40 Proteins in Saccharomyces cerevisiae and Their Orthologs in Candida albicans.Protein J. 2019 Feb;38(1):58-75. doi: 10.1007/s10930-018-9804-8. Protein J. 2019. PMID: 30511317
-
MTL-independent phenotypic switching in Candida tropicalis and a dual role for Wor1 in regulating switching and filamentation.PLoS Genet. 2013 Mar;9(3):e1003369. doi: 10.1371/journal.pgen.1003369. Epub 2013 Mar 21. PLoS Genet. 2013. PMID: 23555286 Free PMC article.
-
Fungal mating pheromones: choreographing the dating game.Fungal Genet Biol. 2011 Jul;48(7):668-76. doi: 10.1016/j.fgb.2011.04.001. Epub 2011 Apr 8. Fungal Genet Biol. 2011. PMID: 21496492 Free PMC article. Review.
-
Deletion of a Yci1 Domain Protein of Candida albicans Allows Homothallic Mating in MTL Heterozygous Cells.mBio. 2016 Apr 26;7(2):e00465-16. doi: 10.1128/mBio.00465-16. mBio. 2016. PMID: 27118591 Free PMC article.
-
The beta subunit of the heterotrimeric G protein triggers the Kluyveromyces lactis pheromone response pathway in the absence of the gamma subunit.Mol Biol Cell. 2010 Feb 1;21(3):489-98. doi: 10.1091/mbc.e09-06-0472. Epub 2009 Dec 16. Mol Biol Cell. 2010. PMID: 20016006 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
