

Auditory response properties of neurons in the tectal longitudinal column of the rat
- PMID: 18662764
- PMCID: PMC2635887
- DOI: 10.1016/j.heares.2008.07.001
Auditory response properties of neurons in the tectal longitudinal column of the rat
Abstract
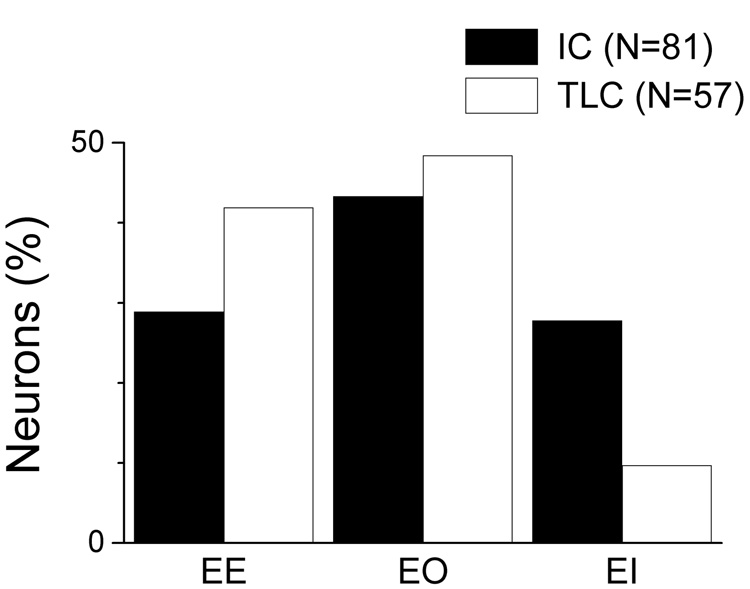
The newly-discovered tectal longitudinal column (TLC) spans the paramedian region of the mammalian tectum. It has connections with several nuclei of the auditory system. In this report, we provide the first detailed description of the responses of TLC neurons to auditory stimuli, including monaural and binaural tones and amplitude modulated tones. For comparison, responses in the inferior colliculus (IC) were also recorded. Neurons in the TLC were sensitive to similar ranges of frequency as IC neurons, could have comparably low thresholds, and showed primarily excitatory responses to stimulation of the contralateral ear with either phasic or sustained response patterns. Differences of TLC compared to IC neurons included broader frequency tuning, higher average threshold, longer response latencies, little synchronization or rate tuning to amplitude modulation frequency and a smaller degree of inhibition evoked by stimulation of the ipsilateral ear. These features of TLC neurons suggest a role for the TLC in descending auditory pathways.
Figures








References
-
- Bock GR, Steel KP. Use of albino animals for auditory research. Hear Res. 1984;13:201–202. - PubMed
-
- Chebat DR, Boire D, Ptito M. Development of the commissure of the superior colliculus in the hamster. J Comp Neurol. 2006;494:887–902. - PubMed
-
- Doron NN, Ledoux JE, Semple MN. Redefining the tonotopic core of rat auditory cortex: physiological evidence for a posterior field. J Comp Neurol. 2002;453:345–360. - PubMed
-
- Faye-Lund H. Projection from the inferior colliculus to the superior olivary complex in the albino rat. Anat Embryol (Berl) 1986;175:35–52. - PubMed
-
- Feliciano M, Saldaña E, Mugnaini A. Direct projections from the rat primary auditory neocortex to nucleus sagulum, paralemniscal regions, superior olivary complex and cochlea nuclei. Auditory Neurosci. 1995;1:287–308.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
