

Expression patterns of transcribed human endogenous retrovirus HERV-K(HML-2) loci in human tissues and the need for a HERV Transcriptome Project
- PMID: 18664271
- PMCID: PMC2525661
- DOI: 10.1186/1471-2164-9-354
Expression patterns of transcribed human endogenous retrovirus HERV-K(HML-2) loci in human tissues and the need for a HERV Transcriptome Project
Abstract
Background: A significant proportion of the human genome is comprised of human endogenous retroviruses (HERVs). HERV transcripts are found in every human tissue. Expression of proviruses of the HERV-K(HML-2) family has been associated with development of human tumors, in particular germ cell tumors (GCT). Very little is known about transcriptional activity of individual HML-2 loci in human tissues, though.
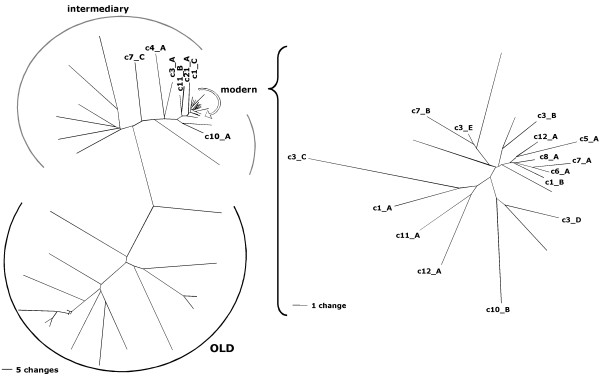
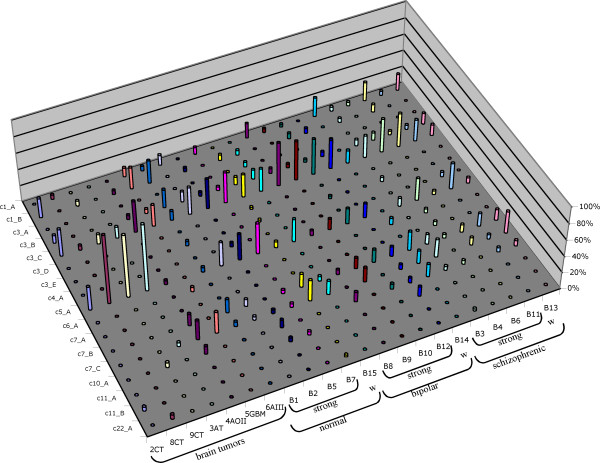
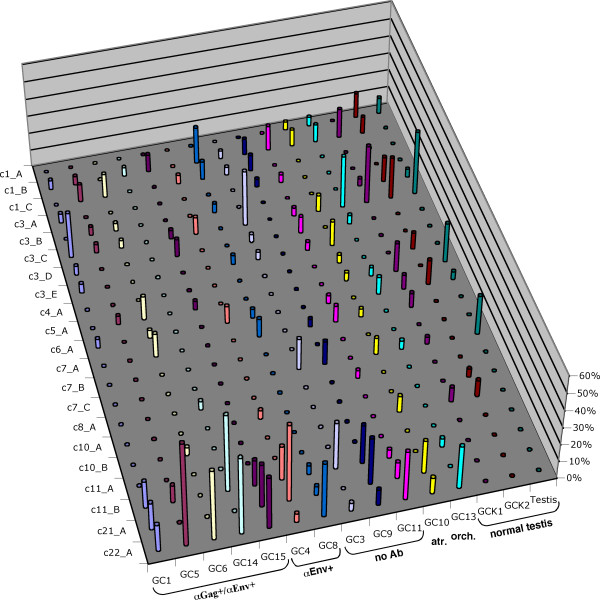
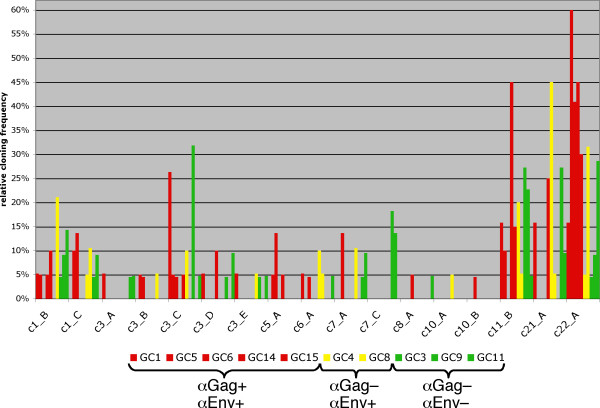
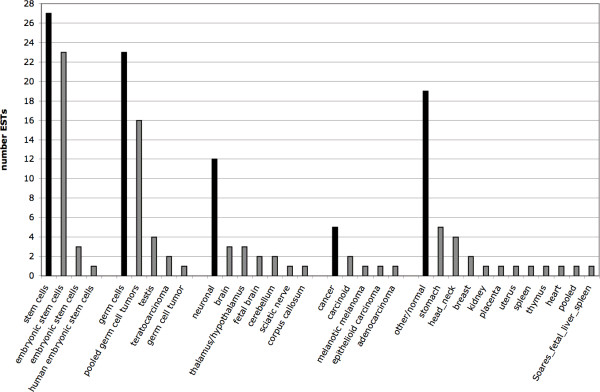
Results: By employing private nucleotide differences between loci, we assigned approximately 1500 HML-2 cDNAs to individual HML-2 loci, identifying, in total, 23 transcriptionally active HML-2 proviruses. Several loci are active in various human tissue types. Transcription levels of some HML-2 loci appear higher than those of other loci. Several HML-2 Rec-encoding loci are expressed in GCT and non-GCT tissues. A provirus on chromosome 22q11.21 appears strongly upregulated in pathologic GCT tissues and may explain high HML-2 Gag protein levels in GCTs. Presence of Gag and Env antibodies in GCT patients is not correlated with activation of individual loci. HML-2 proviruses previously reported capable of forming an infectious HML-2 variant are transcriptionally active in germ cell tissue. Our study furthermore shows that Expressed Sequence Tag (EST) data are insufficient to describe transcriptional activity of HML-2 and other HERV loci in tissues of interest.
Conclusion: Our, to date, largest-scale study reveals in greater detail expression patterns of individual HML-2 loci in human tissues of clinical interest. Moreover, large-scale, specialized studies are indicated to better comprehend transcriptional activity and regulation of HERVs. We thus emphasize the need for a specialized HERV Transcriptome Project.
Figures
References
-
- Mayer J, Meese E. Human endogenous retroviruses in the primate lineage and their influence on host genomes. Cytogenet Genome Res. 2005;110:448–456. - PubMed
-
- de Parseval N, Heidmann T. Human endogenous retroviruses: from infectious elements to human genes. Cytogenet Genome Res. 2005;110:318–332. - PubMed
-
- Sverdlov ED. Perpetually mobile footprints of ancient infections in human genome. FEBS Lett. 1998;428:1–6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
