

A pH-stabilizing role of voltage-gated proton channels in IgE-mediated activation of human basophils
- PMID: 18664579
- PMCID: PMC2504794
- DOI: 10.1073/pnas.0800886105
A pH-stabilizing role of voltage-gated proton channels in IgE-mediated activation of human basophils
Abstract
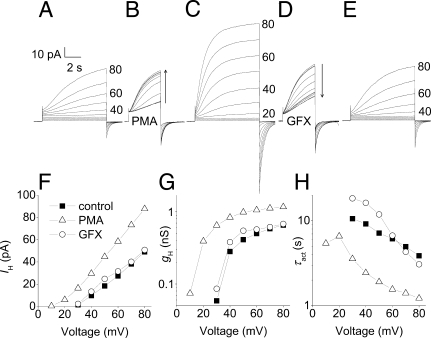
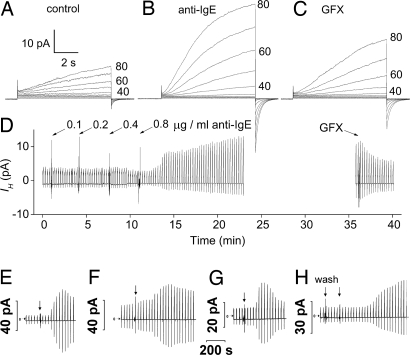
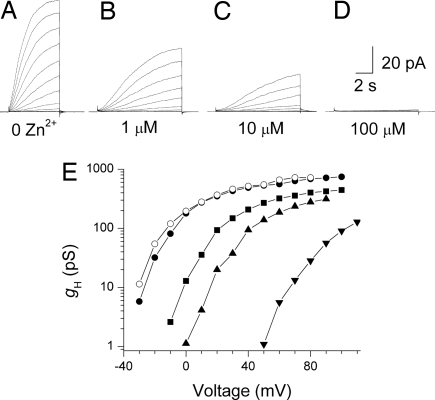
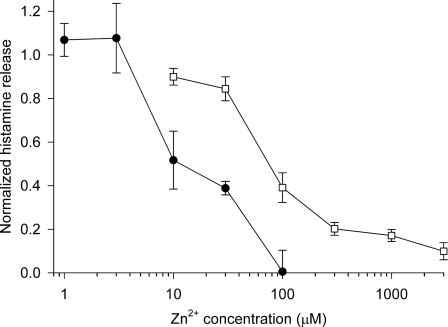
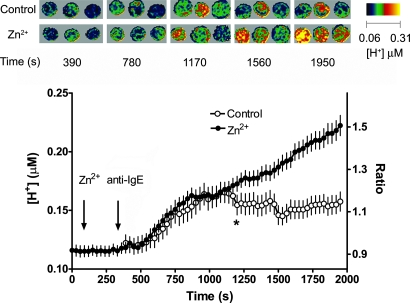
Eosinophils and other phagocytes use NADPH oxidase to kill bacteria. Proton channels in human eosinophils and neutrophils are thought to sustain NADPH oxidase activity, and their opening is greatly enhanced by a variety of NADPH oxidase activators, including phorbol myristate acetate (PMA). In nonphagocytic cells that lack NADPH oxidase, no clear effect of PMA on proton channels has been reported. The basophil is a granulocyte that is developmentally closely related to the eosinophil but nevertheless does not express NADPH oxidase. Thus, one might expect that stimulating basophils with PMA would not affect proton currents. However, stimulation of human basophils in perforated-patch configuration with PMA, N-formyl-methionyl-leucyl-phenylalanine, or anti-IgE greatly enhanced proton currents, the latter suggesting involvement of proton channels during activation of basophils by allergens through their highly expressed IgE receptor (Fc epsilonRI). The anti-IgE-stimulated response occurred in a fraction of cells that varied among donors and was less profound than that to PMA. PKC inhibition reversed the activation of proton channels, and the proton channel response to anti-IgE or PMA persisted in Ca(2+)-free solutions. Zn(2+) at concentrations that inhibit proton current inhibited histamine release elicited by PMA or anti-IgE. Studied with confocal microscopy by using SNARF-AM and the shifted excitation and emission ratioing of fluorescence approach, anti-IgE produced acidification that was exacerbated in the presence of 100 microM Zn(2+). Evidently, proton channels are active in basophils during IgE-mediated responses and prevent excessive acidification, which may account for their role in histamine release.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Cherny VV, Thomas LL, DeCoursey TE. Voltage-gated proton currents in human basophils. Biol Membr. 2001;18:458–465.
-
- DeCoursey TE. Voltage-gated proton channels and other proton transfer pathways. Physiol Rev. 2003;83:475–579. - PubMed
-
- DeCoursey TE, Morgan D, Cherny VV. The voltage dependence of NADPH oxidase reveals why phagocytes need proton channels. Nature. 2003;422:531–534. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
