

Habituation to repeated stress: get used to it
- PMID: 18667167
- PMCID: PMC2773683
- DOI: 10.1016/j.nlm.2008.07.001
Habituation to repeated stress: get used to it
Abstract
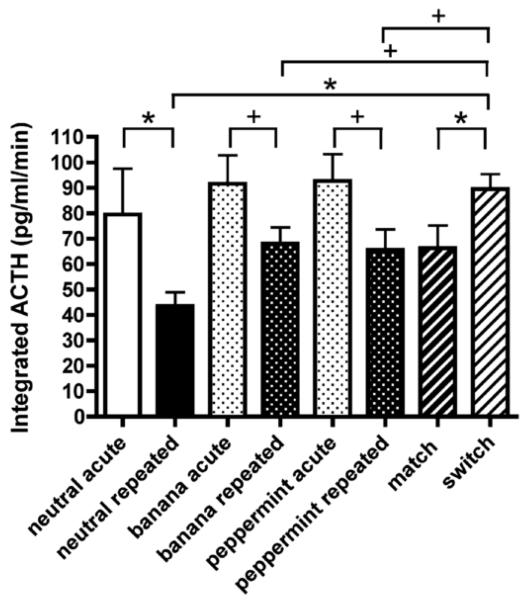
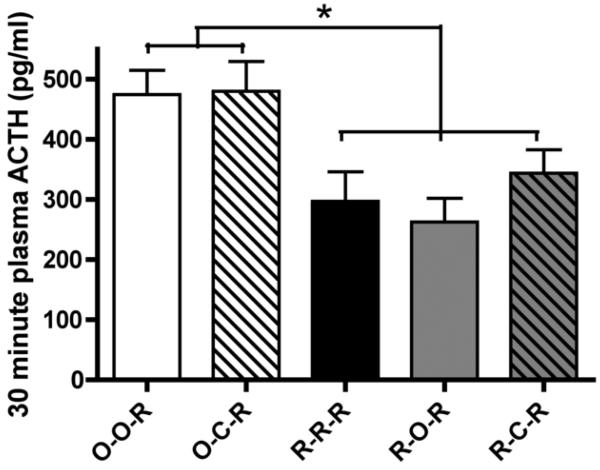
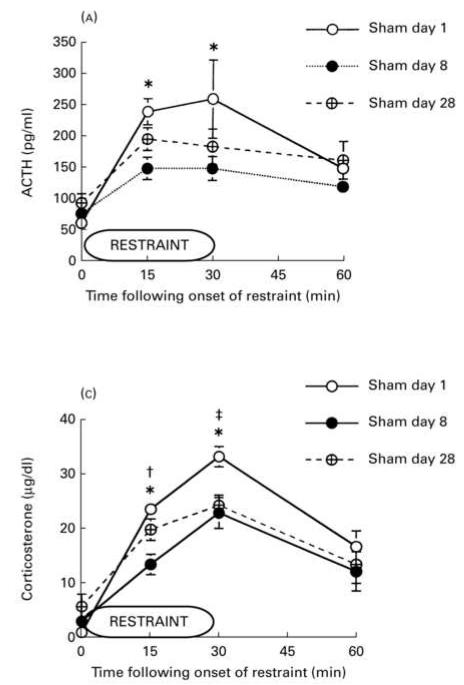
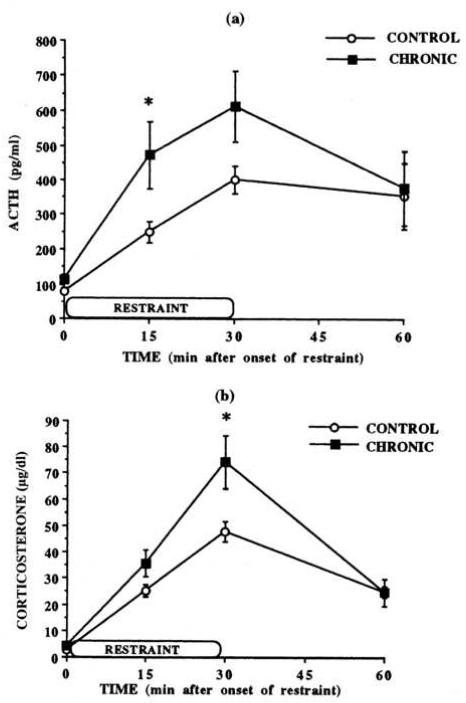
Habituation, as described in the landmark paper by Thompson et al. [Thompson, R. F., & Spencer, W. A. (1966). Habituation: A model phenomenon for the study of neuronal substrates of behavior. Psychological Review, 73(1), 16-43], is a form of simple, nonassociative learning in which the magnitude of the response to a specific stimulus decreases with repeated exposure to that stimulus. A variety of neuronal and behavioral responses have been shown to be subject to habituation based on the criteria presented in that paper. It has been known for several decades that the magnitude of hypothalamic-pituitary-adrenal (HPA) activation occurring in response to a stressor declines with repeated exposure to that same stressor. For some time this decline has been referred to as "habituation" in the stress neurobiology literature. However, how this usage compares to the definition proposed by Thompson and Spencer has not been systematically addressed. For this special issue, we review the stress neurobiology literature and examine the support available for considering declines in HPA response to repeated stress to be response habituation in the sense defined by Thompson and Spencer. We conclude that habituation of HPA activity meets many, but not all, important criteria for response habituation, supporting the use of this term within the context of repeated stress. However, we also propose that response habituation can, at best, only partially explain the phenomenon of HPA habituation, which also involves well-known negative feedback mechanisms, activation of broad stress-related neural circuitry and potentially more complex associative learning mechanisms.
Figures
References
-
- Akana SF, Hanson ES, Horsley CJ, Strack AM, Bhatnagar S, Bradbury MJ, Milligan ED, Dallman MF. Clamped corticosterone (B) reveals the effect of endogenous B on both facilitated responsivity to acute restraint and metabolic responses to chronic stress. Stress (Amsterdam, Netherlands) 1996;1(1):33–49. - PubMed
-
- Akana SF, Shinsako J, Dallman MF. Relationships among adrenal weight, corticosterone, and stimulated adrenocorticotropin levels in rats. Endocrinology. 1983;113(6):2226–2231. - PubMed
-
- Armario A. The hypothalamic-pituitary-adrenal axis: What can it tell us about stressors? CNS & Neurological Disorders Drug Targets. 2006;5(5):485–501. - PubMed
-
- Armario A, Castellanos JM, Balasch J. Adaptation of anterior pituitary hormones to chronic noise stress in male rats. Behavioral and Neural Biology. 1984;41(1):71–76. - PubMed
-
- Armario A, Hidalgo J, Giralt M. Evidence that the pituitary-adrenal axis does not cross-adapt to stressors: Comparison to other physiological variables. Neuroendocrinology. 1988;47(3):263–267. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
