

Microtubules have opposite orientation in axons and dendrites of Drosophila neurons
- PMID: 18667536
- PMCID: PMC2555934
- DOI: 10.1091/mbc.e07-10-1079
Microtubules have opposite orientation in axons and dendrites of Drosophila neurons
Abstract
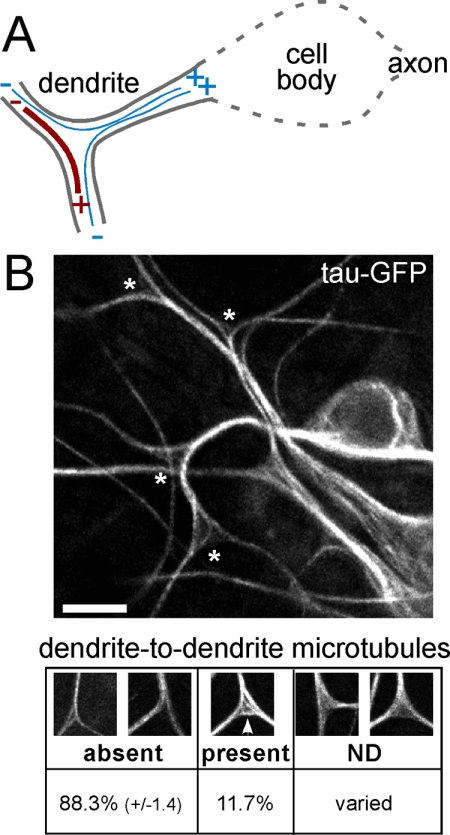
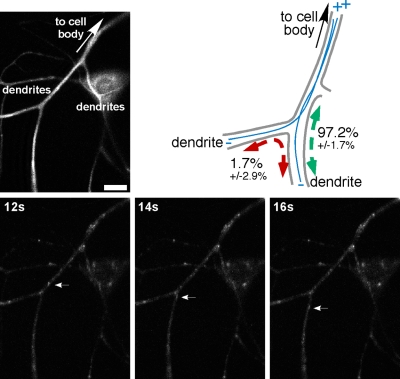
In vertebrate neurons, axons have a uniform arrangement of microtubules with plus ends distal to the cell body (plus-end-out), and dendrites have equal numbers of plus- and minus-end-out microtubules. To determine whether microtubule orientation is a conserved feature of axons and dendrites, we analyzed microtubule orientation in invertebrate neurons. Using microtubule plus end dynamics, we mapped microtubule orientation in Drosophila sensory neurons, interneurons, and motor neurons. As expected, all axonal microtubules have plus-end-out orientation. However, in proximal dendrites of all classes of neuron, approximately 90% of dendritic microtubules were oriented with minus ends distal to the cell body. This result suggests that minus-end-out, rather than mixed orientation, microtubules are the signature of the dendritic microtubule cytoskeleton. Surprisingly, our map of microtubule orientation predicts that there are no tracks for direct cargo transport between the cell body and dendrites in unipolar neurons. We confirm this prediction, and validate the completeness of our map, by imaging endosome movements in motor neurons. As predicted by our map, endosomes travel smoothly between the cell body and axon, but they cannot move directly between the cell body and dendrites.
Figures





Similar articles
-
Dendrites In Vitro and In Vivo Contain Microtubules of Opposite Polarity and Axon Formation Correlates with Uniform Plus-End-Out Microtubule Orientation.J Neurosci. 2016 Jan 27;36(4):1071-85. doi: 10.1523/JNEUROSCI.2430-15.2016. J Neurosci. 2016. PMID: 26818498 Free PMC article.
-
Dynein is required for polarized dendritic transport and uniform microtubule orientation in axons.Nat Cell Biol. 2008 Oct;10(10):1172-80. doi: 10.1038/ncb1777. Epub 2008 Aug 31. Nat Cell Biol. 2008. PMID: 18758451 Free PMC article.
-
Unrestrained growth of correctly oriented microtubules instructs axonal microtubule orientation.Elife. 2022 Oct 10;11:e77608. doi: 10.7554/eLife.77608. Elife. 2022. PMID: 36214669 Free PMC article.
-
Principles of microtubule polarity in linear cells.Dev Biol. 2022 Mar;483:112-117. doi: 10.1016/j.ydbio.2022.01.004. Epub 2022 Jan 8. Dev Biol. 2022. PMID: 35016908 Free PMC article. Review.
-
The role of motor proteins in establishing the microtubule arrays of axons and dendrites.J Chem Neuroanat. 1998 Jun;14(3-4):175-80. doi: 10.1016/s0891-0618(98)00012-x. J Chem Neuroanat. 1998. PMID: 9704896 Review.
Cited by
-
Recent progress in dendritic pruning of Drosophila C4da sensory neurons.Open Biol. 2024 Jul;14(7):240059. doi: 10.1098/rsob.240059. Epub 2024 Jul 24. Open Biol. 2024. PMID: 39046196 Free PMC article. Review.
-
The functions of kinesin and kinesin-related proteins in eukaryotes.Cell Adh Migr. 2020 Dec;14(1):139-152. doi: 10.1080/19336918.2020.1810939. Cell Adh Migr. 2020. PMID: 32842864 Free PMC article. Review.
-
Patronin-mediated minus end growth is required for dendritic microtubule polarity.J Cell Biol. 2019 Jul 1;218(7):2309-2328. doi: 10.1083/jcb.201810155. Epub 2019 May 10. J Cell Biol. 2019. PMID: 31076454 Free PMC article.
-
Building Blocks of Functioning Brain: Cytoskeletal Dynamics in Neuronal Development.Int Rev Cell Mol Biol. 2016;322:183-245. doi: 10.1016/bs.ircmb.2015.10.002. Epub 2016 Jan 6. Int Rev Cell Mol Biol. 2016. PMID: 26940519 Free PMC article. Review.
-
Calcium plays an essential role in early-stage dendrite injury detection and regeneration.Prog Neurobiol. 2024 Aug;239:102635. doi: 10.1016/j.pneurobio.2024.102635. Epub 2024 May 31. Prog Neurobiol. 2024. PMID: 38825174 Free PMC article.
References
-
- Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. Molecular Biology of the Cell. New York: Garland Science; 2002.
-
- Black M. M., Baas P. W. The basis of polarity in neurons. Trends Neurosci. 1989;12:211–214. - PubMed
-
- Burack M. A., Silverman M. A., Banker G. The role of selective transport in neuronal protein sorting. Neuron. 2000;26:465–472. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
