

Peeking into pit fields: a multiple twinning model of secondary plasmodesmata formation in tobacco
- PMID: 18667640
- PMCID: PMC2483367
- DOI: 10.1105/tpc.107.056903
Peeking into pit fields: a multiple twinning model of secondary plasmodesmata formation in tobacco
Abstract
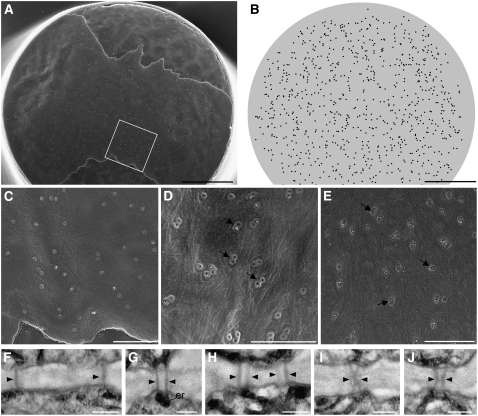
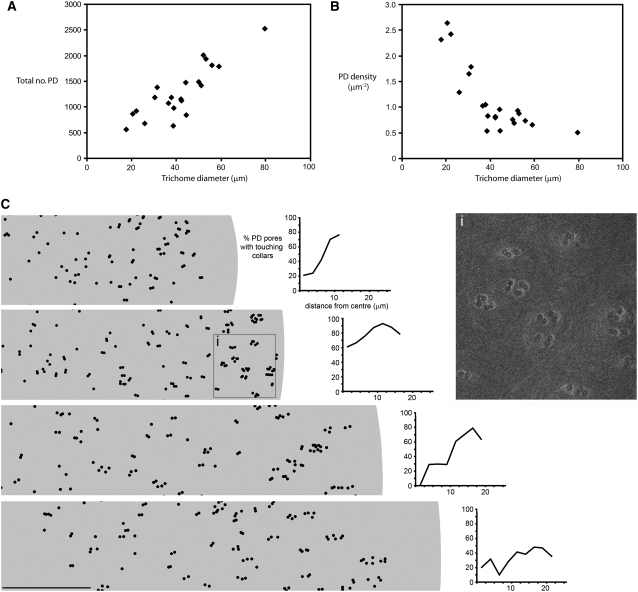
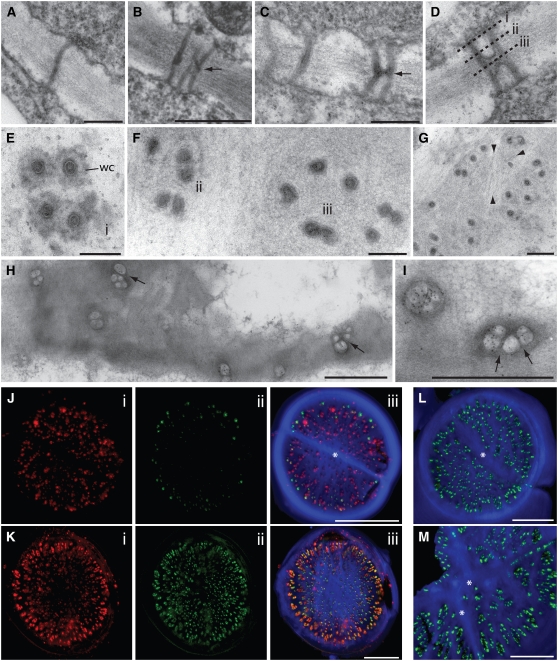
In higher plants, plasmodesmata (PD) are major conduits for cell-cell communication. Primary PD are laid down at cytokinesis, while secondary PD arise during wall extension. During leaf development, the basal cell walls of trichomes extend radially without division, providing a convenient system for studying the origin of secondary PD. We devised a simple freeze-fracture protocol for examining large numbers of PD in surface view. In the postcytokinetic wall, simple PD were distributed randomly. As the wall extended, PD became twinned at the cell periphery. Additional secondary pores were inserted at right angles to these, giving rise to pit fields composed of several paired PD. During wall extension, the number of PD increased fivefold due to the insertion of secondary PD. Our data are consistent with a model in which a subset of the original primary PD pores function as templates for the insertion of new secondary PD, spatially fixing the position of future pit fields. Many of the new PD shared the same wall collar as the original PD pore, suggesting that new PD pores may arise by fissions of existing PD progenitors. Different models of secondary PD formation are discussed. Our data are supported by a computational model, Plasmodesmap, which accurately simulates the formation of radial pit fields during cell wall extension based on the occurrence of multiple PD twinning events in the cell wall. The model predicts PD distributions with striking resemblance to those seen on fractured wall faces.
Figures




References
-
- Angell, S.M., Davies, C., and Baulcombe, D.C. (1996). Cell-to-cell movement of potato virus X is associated with a change in the size exclusion limit of plasmodesmata in trichome cells of Nicotiana clevelandii. Virology 216 197–201. - PubMed
-
- Badelt, K., White, R.G., Overall, R.L., and Vesk, M. (1994). Ultrastructural specialisations of the wall sleeve around plasmodesmata. Am. J. Bot. 81 1422–1477.
-
- Baluska, F., Samaj, J., Napier, R., and Volkmann, D. (1999). Maize calreticulin localizes preferentially to plasmodesmata in root apex. Plant J. 14 481–488. - PubMed
-
- Barnett, J.R. (1987). Changes in the distribution of plasmodesmata in developing fibre-tracheid pit membranes of Sorbus aucuparia L. Ann. Bot. (Lond.) 59 269–279.
-
- Baskin, T. (2005). Anisotropic expansion of the plant cell wall. Annu. Rev. Cell Dev. Biol. 21 203–222. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
