

Light-activated DNA binding in a designed allosteric protein
- PMID: 18667691
- PMCID: PMC2504796
- DOI: 10.1073/pnas.0709610105
Light-activated DNA binding in a designed allosteric protein
Abstract
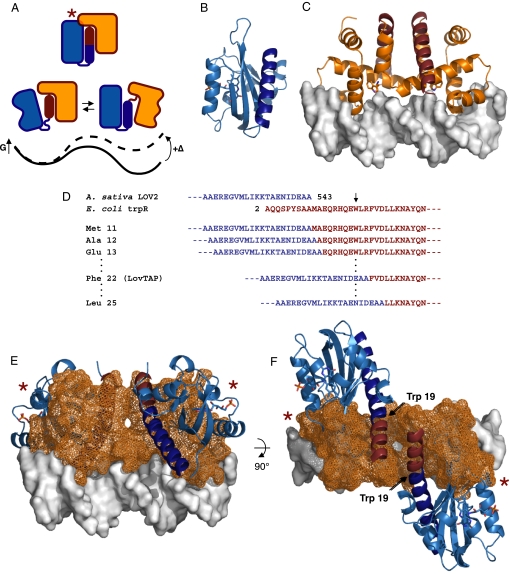
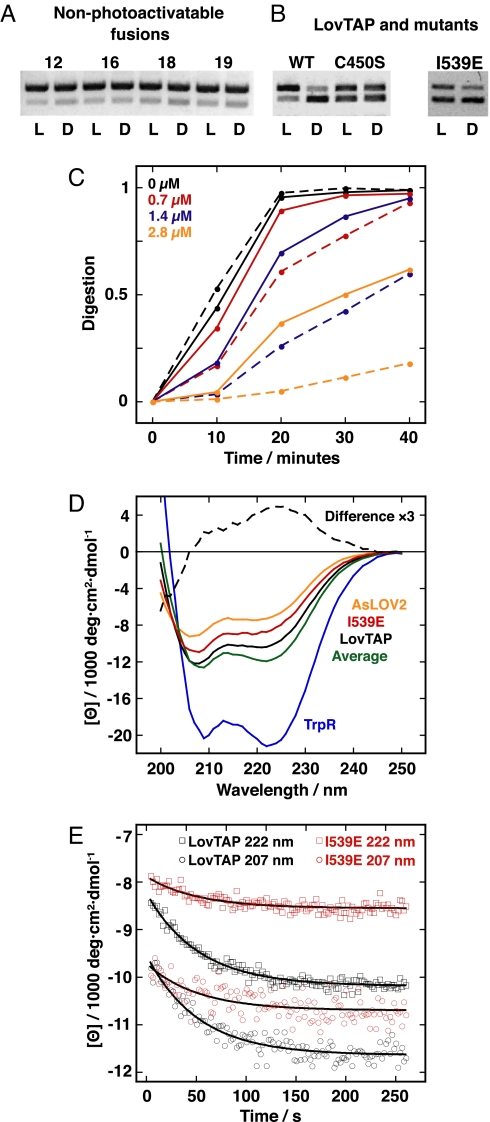
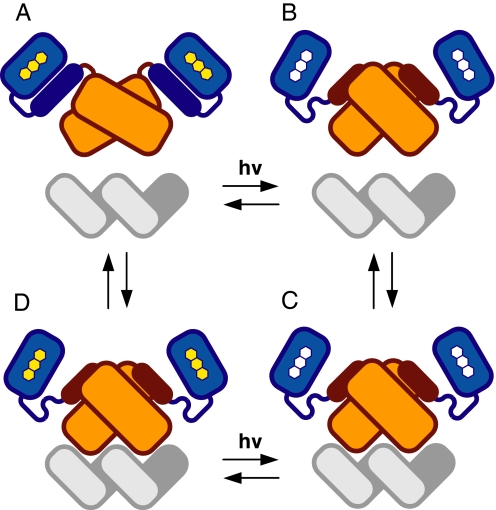
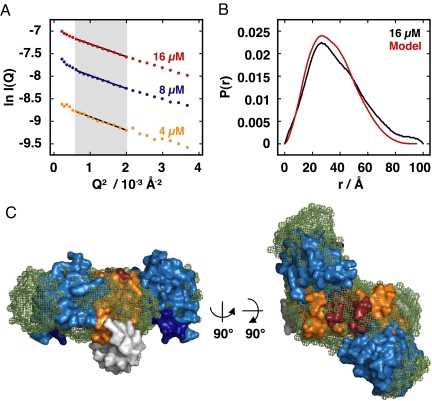
An understanding of how allostery, the conformational coupling of distant functional sites, arises in highly evolvable systems is of considerable interest in areas ranging from cell biology to protein design and signaling networks. We reasoned that the rigidity and defined geometry of an alpha-helical domain linker would make it effective as a conduit for allosteric signals. To test this idea, we rationally designed 12 fusions between the naturally photoactive LOV2 domain from Avena sativa phototropin 1 and the Escherichia coli trp repressor. When illuminated, one of the fusions selectively binds operator DNA and protects it from nuclease digestion. The ready success of our rational design strategy suggests that the helical "allosteric lever arm" is a general scheme for coupling the function of two proteins.
Conflict of interest statement
A patent application relating to this work has been filed for which D.S., K.M., and T.R.S. are coinventors.
Figures
References
-
- Bhattacharyya RP, Remenyi A, Yeh BJ, Lim WA. Domains, motifs, and scaffolds: The role of modular interactions in the evolution and wiring of cell signaling circuits. Annu Rev Biochem. 2006;75:655–680. - PubMed
-
- Pawson T, Linding R. Synthetic modular systems—Reverse engineering of signal transduction. FEBS Lett. 2005;579:1808–1814. - PubMed
-
- Pawson T, Nash P. Assembly of cell regulatory systems through protein interaction domains. Science. 2003;300:445–452. - PubMed
-
- Ponting CP, Russell RR. The natural history of protein domains. Annu Rev Biophys Biomol Struct. 2002;31:45–71. - PubMed
-
- Ostermeier M. Engineering allosteric protein switches by domain insertion. Protein Eng Des Sel. 2005;18:359–364. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
