

The roles of CD8 central and effector memory T-cell subsets in allograft rejection
- PMID: 18671680
- PMCID: PMC4872301
- DOI: 10.1111/j.1600-6143.2008.02335.x
The roles of CD8 central and effector memory T-cell subsets in allograft rejection
Abstract
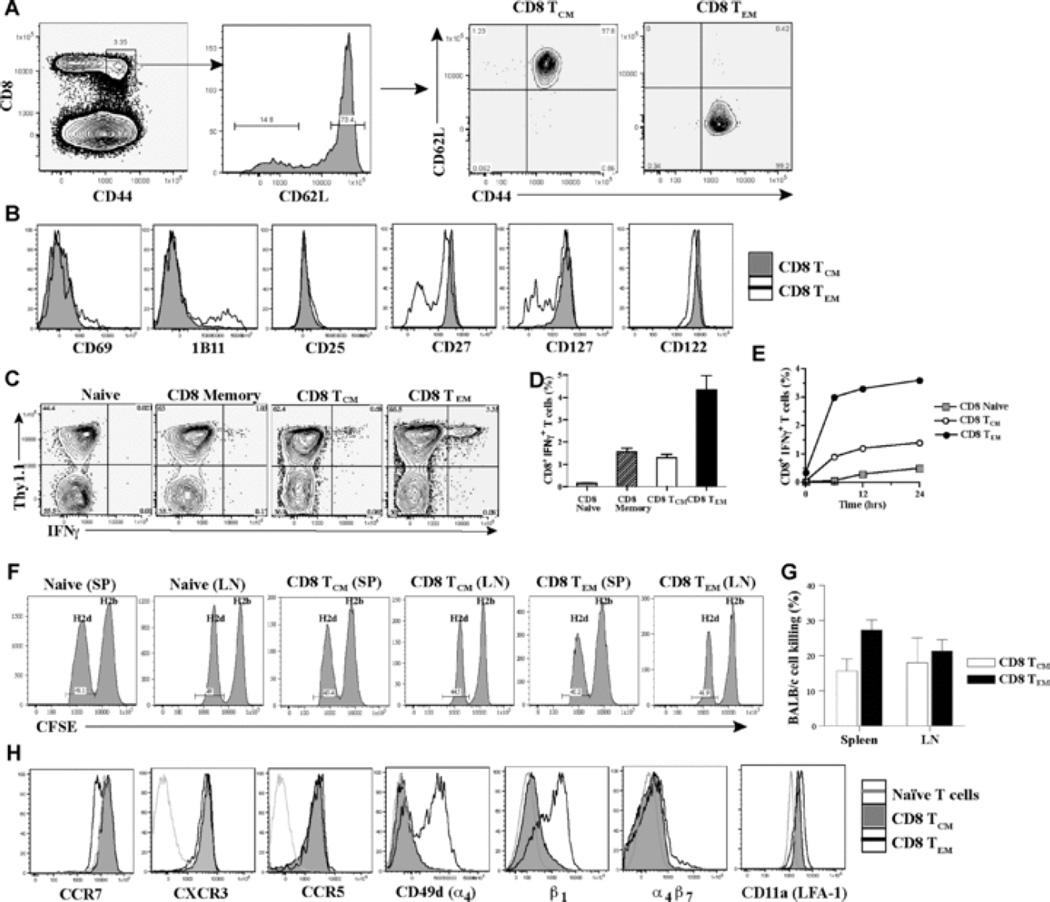
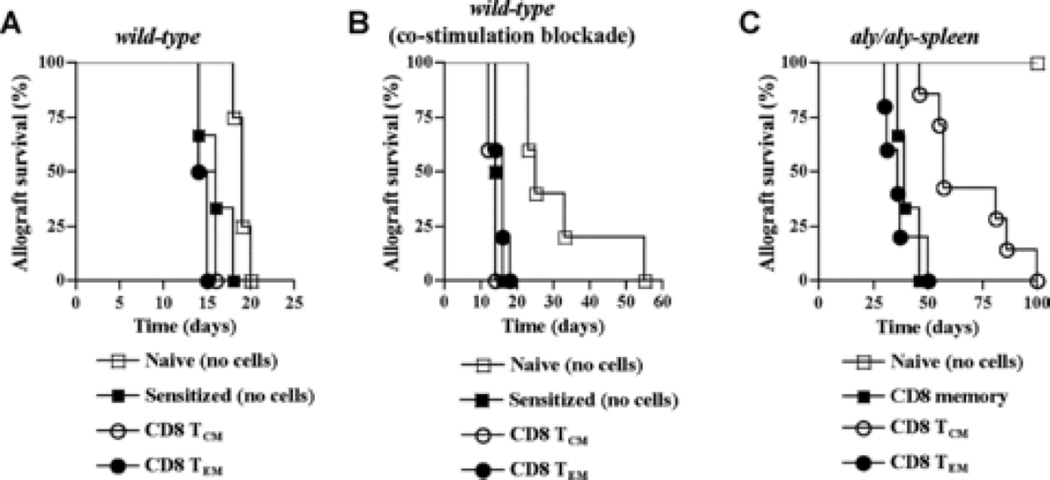
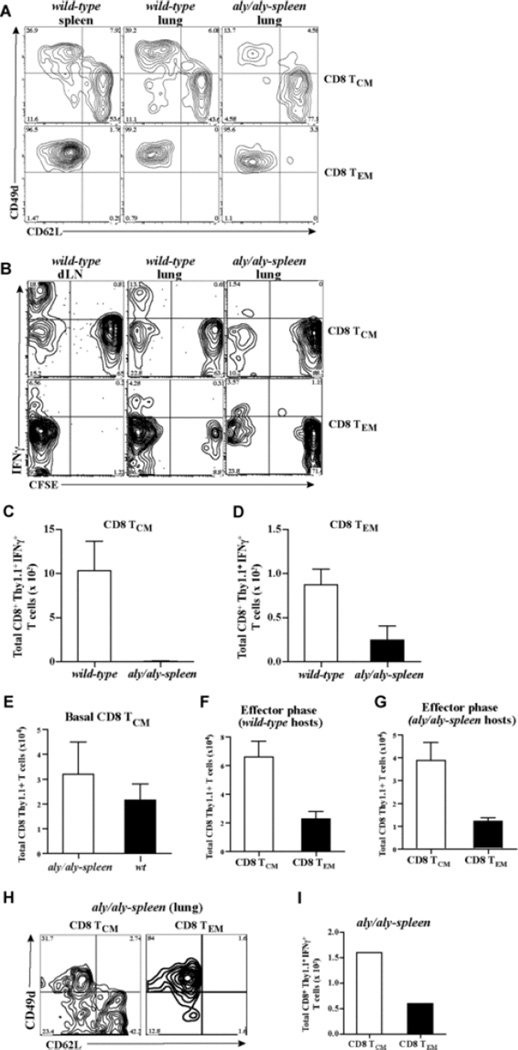
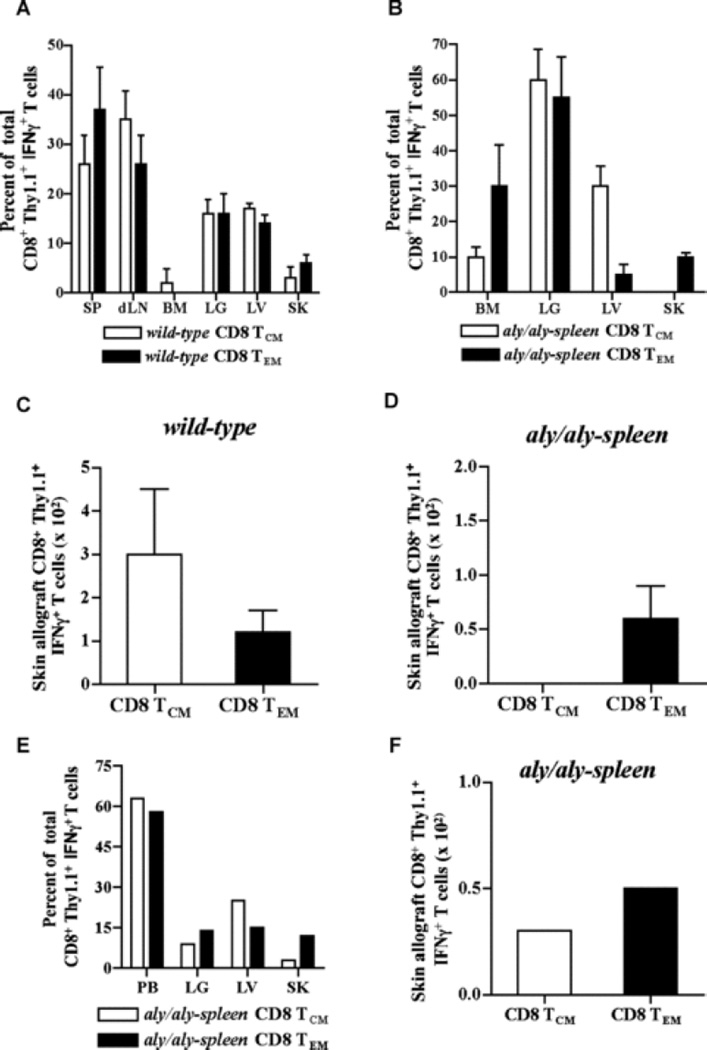
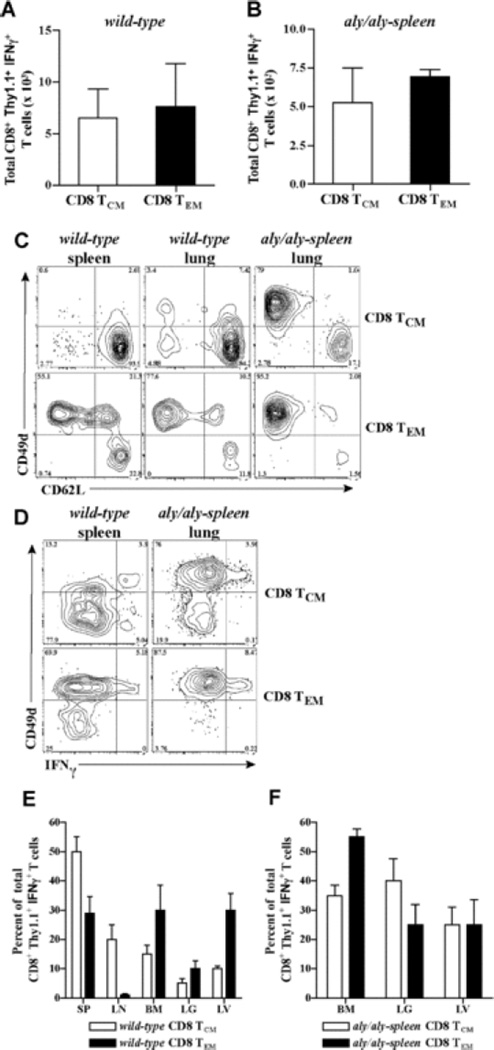
The contribution of secondary lymphoid tissue-homing central memory T cells (T(CM)) and peripheral tissue-homing effector memory T cells (T(EM)) to allograft rejection is not known. We tested whether T(EM) is the principal subset responsible for allograft rejection due to the nonlymphoid location of target antigens. Skin allograft rejection was studied after transferring either CD8 T(CM) or T(EM) to wild-type mice and to mice that lack secondary lymphoid tissues. We found that CD8 T(CM) and T(EM) were equally effective at rejecting allografts in wild-type hosts. However, CD8 T(EM) were significantly better than T(CM) at rejecting allografts in the absence of secondary lymphoid tissues. CD8 T(CM) were dependent upon secondary lymphoid tissues more than T(EM) for optimal differentiation into effectors that migrate into the allograft. Recall of either CD8 T(CM) or T(EM) led to accumulation of T(EM) after allograft rejection. These findings indicate that either CD8 T(CM) or T(EM) mediate allograft rejection but T(EM) have an advantage over T(CM) in immune surveillance of peripheral tissues, including transplanted organs.
Figures
References
-
- Heeger PS, Greenspan NS, Kuhlenschmidt S, Dejelo C, Hricik DE, Schulak JA, et al. Pretransplant frequency of donor-specific, IFN-γ-producing lymphocytes is a manifestation of immunologic memory and correlates with the risk of posttransplant rejection episodes. J Immunol. 1999;163:2267–2275. - PubMed
-
- Najafian N, Salama A, Fedoseyeva E, Benichou G, Sayegh M. Enzyme-linked immunosorbent spot assay analysis of peripheral blood lymphocyte reactivity to donor HLA-DR peptides: potential novel assay for prediction of outcomes for renal transplant recipients. J Am Soc Nephrol. 2002;13:252–259. - PubMed
-
- Pearl JP, Parris J, Hale DA, Hoffman SC, Bernstein WB, McCoy KL, et al. Immunocompetent T cells with a memory-like phenotype are the dominant cell type following antibody-mediated T-cell depletion. Am J Transplant. 2005;5:465–474. - PubMed
-
- Valujskikh A, Pantenburg B, Heeger P. Primed allospecific T cells prevent the effects of costimulatory blockade on prolonged cardiac allograft survival in mice. Am J Transplant. 2002;2:501–509. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
