

Dynamic partitioning of mitotic kinesin-5 cross-linkers between microtubule-bound and freely diffusing states
- PMID: 18678711
- PMCID: PMC2500124
- DOI: 10.1083/jcb.200804100
Dynamic partitioning of mitotic kinesin-5 cross-linkers between microtubule-bound and freely diffusing states
Abstract
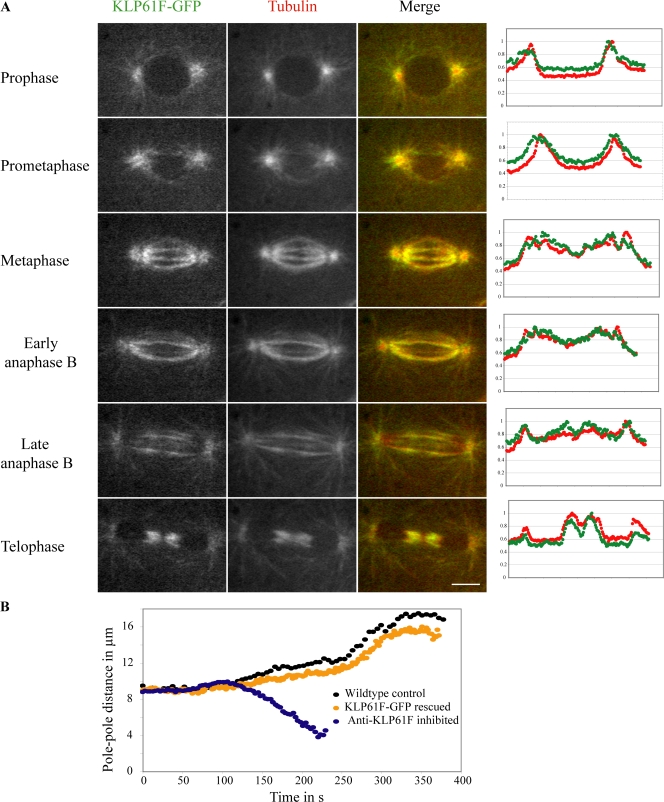
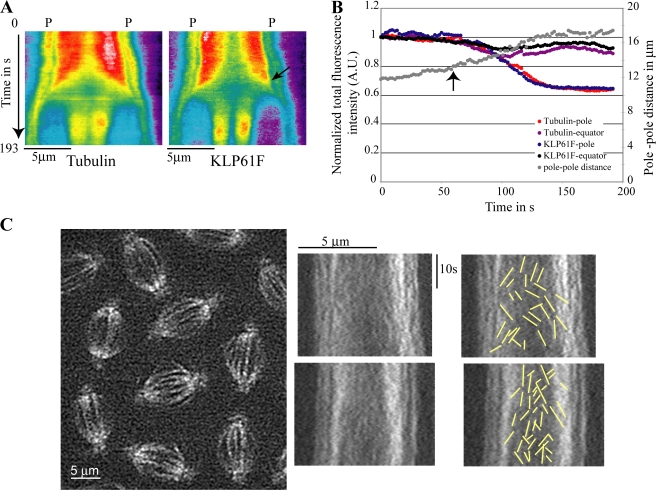
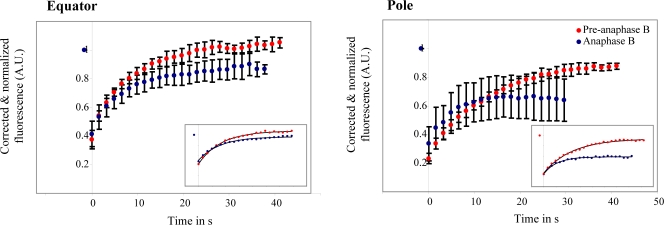
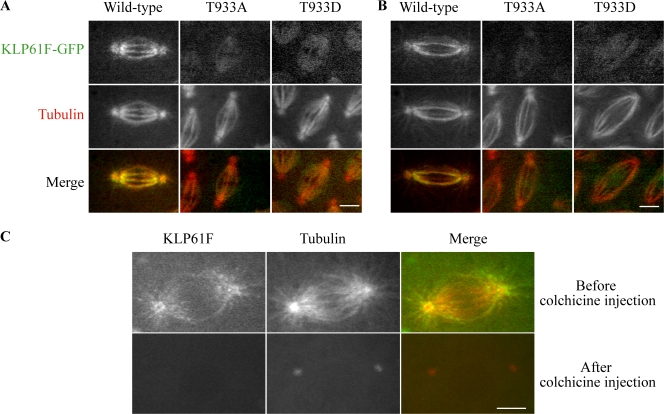
The dynamic behavior of homotetrameric kinesin-5 during mitosis is poorly understood. Kinesin-5 may function only by binding, cross-linking, and sliding adjacent spindle microtubules (MTs), or, alternatively, it may bind to a stable "spindle matrix" to generate mitotic movements. We created transgenic Drosophila melanogaster expressing fluorescent kinesin-5, KLP61F-GFP, in a klp61f mutant background, where it rescues mitosis and viability. KLP61F-GFP localizes to interpolar MT bundles, half spindles, and asters, and is enriched around spindle poles. In fluorescence recovery after photobleaching experiments, KLP61F-GFP displays dynamic mobility similar to tubulin, which is inconsistent with a substantial static pool of kinesin-5. The data conform to a reaction-diffusion model in which most KLP61F is bound to spindle MTs, with the remainder diffusing freely. KLP61F appears to transiently bind MTs, moving short distances along them before detaching. Thus, kinesin-5 motors can function by cross-linking and sliding adjacent spindle MTs without the need for a static spindle matrix.
Figures
Similar articles
-
Kinesin-5 in Drosophila embryo mitosis: sliding filament or spindle matrix mechanism?Cell Motil Cytoskeleton. 2009 Aug;66(8):500-8. doi: 10.1002/cm.20349. Cell Motil Cytoskeleton. 2009. PMID: 19291760 Free PMC article.
-
The bipolar kinesin, KLP61F, cross-links microtubules within interpolar microtubule bundles of Drosophila embryonic mitotic spindles.J Cell Biol. 1999 Jan 11;144(1):125-38. doi: 10.1083/jcb.144.1.125. J Cell Biol. 1999. PMID: 9885249 Free PMC article.
-
Kinesin-5-dependent poleward flux and spindle length control in Drosophila embryo mitosis.Mol Biol Cell. 2009 Mar;20(6):1749-62. doi: 10.1091/mbc.e08-10-1033. Epub 2009 Jan 21. Mol Biol Cell. 2009. PMID: 19158379 Free PMC article.
-
Mitotic motors and chromosome segregation: the mechanism of anaphase B.Biochem Soc Trans. 2011 Oct;39(5):1149-53. doi: 10.1042/BST0391149. Biochem Soc Trans. 2011. PMID: 21936780 Review.
-
Mechanisms of Mitotic Spindle Assembly.Annu Rev Biochem. 2016 Jun 2;85:659-83. doi: 10.1146/annurev-biochem-060815-014528. Epub 2016 Apr 21. Annu Rev Biochem. 2016. PMID: 27145846 Free PMC article. Review.
Cited by
-
Towards a quantitative understanding of mitotic spindle assembly and mechanics.J Cell Sci. 2010 Oct 15;123(Pt 20):3435-45. doi: 10.1242/jcs.062208. J Cell Sci. 2010. PMID: 20930139 Free PMC article. Review.
-
Mitotic force generators and chromosome segregation.Cell Mol Life Sci. 2010 Jul;67(13):2231-50. doi: 10.1007/s00018-010-0326-6. Epub 2010 Mar 10. Cell Mol Life Sci. 2010. PMID: 20221784 Free PMC article. Review.
-
Kinesin-5: cross-bridging mechanism to targeted clinical therapy.Gene. 2013 Dec 1;531(2):133-49. doi: 10.1016/j.gene.2013.08.004. Epub 2013 Aug 14. Gene. 2013. PMID: 23954229 Free PMC article. Review.
-
Kinesin-5 in Drosophila embryo mitosis: sliding filament or spindle matrix mechanism?Cell Motil Cytoskeleton. 2009 Aug;66(8):500-8. doi: 10.1002/cm.20349. Cell Motil Cytoskeleton. 2009. PMID: 19291760 Free PMC article.
-
Astral microtubule cross-linking safeguards uniform nuclear distribution in the Drosophila syncytium.J Cell Biol. 2022 Jan 3;221(1):e202007209. doi: 10.1083/jcb.202007209. Epub 2021 Nov 12. J Cell Biol. 2022. PMID: 34766978 Free PMC article.
References
-
- Ashburner, M. 1989. Drosophila: A Laboratory Handbook. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 1,331 pp.
-
- Brust-Mascher, I., and J.M. Scholey. 2007. Mitotic spindle dynamics in Drosophila. Int. Rev. Cytol. 259:139–172. - PubMed
-
- Bulinski, J.C., D.J. Odde, B.J. Howell, T.D. Salmon, and C.M. Waterman-Storer. 2001. Rapid dynamics of the microtubule binding of ensconsin in vivo. J. Cell Sci. 114:3885–3897. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
