

Review
doi: 10.1104/pp.108.121756.
PIN polar targeting
Affiliations
- PMID: 18678746
- PMCID: PMC2492634
- DOI: 10.1104/pp.108.121756
Item in Clipboard
Review
PIN polar targeting
Plant Physiol.
2008 Aug.
No abstract available
Figures
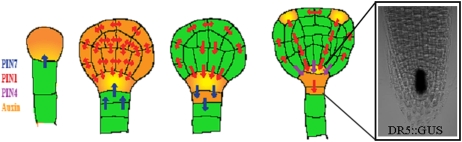
Establishment of apical-basal polarity during Arabidopsis embryogenesis. At early stages, PIN7 localizes to the apical sides of suspensor cells mediating auxin flow into the proembryo, where PIN1 is localized first in a nonpolar manner. At the middle globular stage, PIN1 basal polarity in provascular cells is established, followed by PIN7 relocation to the basal side of suspensor cells. These PIN polarity rearrangements result in redirection of the auxin flow to the basal part of the embryo, where auxin accumulation contributes to root meristem specification. At later stages, PIN4 expression in the central root meristem aids the establishment of local auxin accumulation in the center of developing root meristems. Furthermore, PIN1 relocates at the apical surface of the proembryo to establish two symmetrically positioned auxin accumulation foci marking sites of future embryonic leaves.
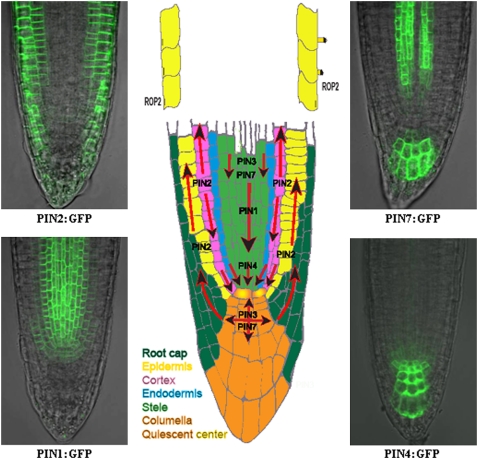
Polar localization of PIN proteins in the Arabidopsis root tip. The directionality of auxin transport (arrows) is determined by the polar, subcellular localization of PIN proteins. PIN1 is localized at the basal (root apex-facing) side of the root vasculature; PIN2 at the basal side of the cortical cells and at the apical (shoot apex-facing) side of the epidermal and root cap cells; PIN3 in an apolar manner in the columella cells of the root; PIN4 at the basal side of cells in the central root meristem and with less pronounced polarity in the cells of the quiescent center; and PIN7 at the basal side of the stele cells and apolar in the columella cells. ROP2 is also asymmetrically localized, associated with the places of root hair formation. Examples of PIN1:GFP, PIN2:GFP, PIN4:GFP, and PIN7:GFP expression in the root are depicted.
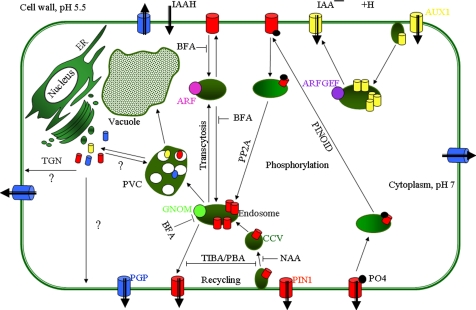
Overview of the subcellular trafficking routes in a polarized plant cell. Auxin influx and efflux (black thick arrows) are mediated by AUX1 and PIN proteins, respectively. PGP ATP-binding cassette-type transporters are also involved in auxin efflux. According to the chemiosmotic model, auxin in the protonated form can also enter the cell passively. Basally localized PIN1 recycles in a BFA-sensitive and GNOM-dependent manner between endosomes and the PM. Constitutive cycling of vesicles between these two compartments is disrupted by 2,3,5-triiodobenzoic acid (TIBA) and 1-pyrenoylbenzoic acid (PBA), drugs that inhibit auxin transport and actin cytoskeleton dynamics (Dhonukshe et al., 2008). AUX1 also displays constitutive cycling to and from the apical PM that is dependent of an unknown BFA-insensitive ARF GEF. Auxin, such as α-naphthaleneacetic acid (NAA), inhibits PIN internalization, increasing the amount and activity of PINs at the PM (Paciorek et al., 2005). PID kinase and PP2A phosphatase mediate reversible PIN phosphorylation, thus contributing to the decision on the apical-basal PIN targeting. Phosphorylated and dephosphorylated PIN proteins are preferentially sorted into the apical and basal targeting pathways, respectively (Michniewicz et al., 2007). PIN1 is able to translocate between the basal and apical cell sides by a combination of constitutive endocytic cycling and alternative recruitment by distinct ARF GEF-dependent apical and basal targeting machineries (Kleine-Vehn et al., 2008).
References
-
- Benjamins R, Quint A, Weijers D, Hooykaas P, Offringa R (2001) The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 128 4057–4067 - PubMed
-
- Benková E, Michniewicz M, Sauer M, Teichmann T, Seifertová D, Jürgens G, Friml J (2003) Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115 591–602 - PubMed
-
- Bennett MJ, Marchant A, Green HG, May ST, Ward SP, Millner PA, Walker AR, Schulz B, Feldmann KA (1996) Arabidopsis AUX1 gene: a permease-like regulator of root gravitropism. Science 273 948–950 - PubMed
-
- Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I, Friml J, Heidstra R, Aida M, Palme K, Scheres B (2005) The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 433 39–44 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
