

doi: 10.1104/pp.108.121566.
Secretory pathways in plant immune responses
Affiliations
- PMID: 18678749
- PMCID: PMC2492620
- DOI: 10.1104/pp.108.121566
Item in Clipboard
Secretory pathways in plant immune responses
Plant Physiol.
2008 Aug.
No abstract available
Figures
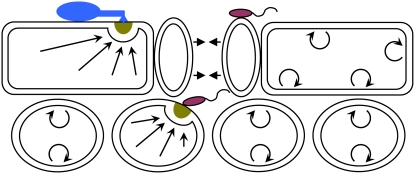
Dual function of secretory pathways in plant immune responses and plant development. Leaf cross section showing epidermal and mesophyll cell layers. Single plant cells target immune responses to contact sites of fungal or bacterial pathogens, leading to papilla formation (dark yellow semicircle) in the paramural space. The indicated flagellate bacteria (red) enter the leaf interior through stomata and remain in the apoplastic space for multiplication. The indicated fungal parasite (blue) attempts to penetrate the plant cell wall to access nutrients from an epidermal cell. Stomatal closure (denoted by short arrows) is one resistance mechanism against bacterial ingress (Melotto et al., 2006). Known vesicle-associated and SNARE protein-mediated exocytosis pathways drive focal (long straight arrows) as well as nondirectional (semicircles with arrowheads) secretion of antimicrobial cargo into the apoplastic space. A second function of these secretory pathways in plant development might involve constitutive nondirectional secretion of cell wall building blocks and cell wall modifying enzymes during cell growth.
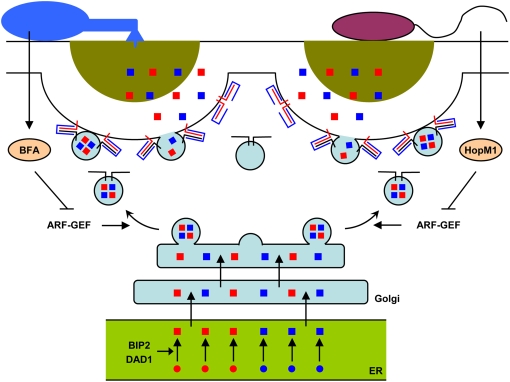
Targeted vesicle-associated and SNARE protein-mediated secretion at microbial contact sites. The indicated vesicles are targeted to the plant plasma membrane beneath a bacterial (right) or fungal (left) contact site along polarized actin cables (not shown). Vesicles are thought to be loaded with cargo derived from the ER/Golgi protein secretory pathway. Vesicles contain constitutive molecules required for the maintenance of plasma membrane and cell wall functions (blue squares) and/or transport additional pathogen-inducible antimicrobial molecules (red squares), leading to the formation of a papillary cell wall scaffold containing toxic cocktails (dark yellow semicircle). At contact sites with powdery mildew fungi (left), vesicle cargo is discharged subsequent to ternary SNARE complex formation involving plasma membrane-resident PEN1 syntaxin (red line), SNAP33 (blue line), and endomembrane-resident VAMP721/722 (black line). At contact sites with bacterial pathogens (right), resistance responses require plasma membrane-resident SYP132 syntaxin (red line), and possibly SNAP33 (blue line) as well as yet unknown VAMPs (black line). During immune responses to bacteria, expansion of the protein-folding machinery including BIP2 and DAD1 is believed to reflect an increased cellular requirement to build up secreted PR proteins in the ER (green). One defense molecule secreted in a SYP132-dependent manner is the pathogen-inducible PR-1 protein. Fungi and bacteria can interfere with the host secretory pathway by inhibiting ARF-GEFs, which regulate vesicle formation, using effector molecules such as brefeldin A (BFA) or HopM1, respectively.
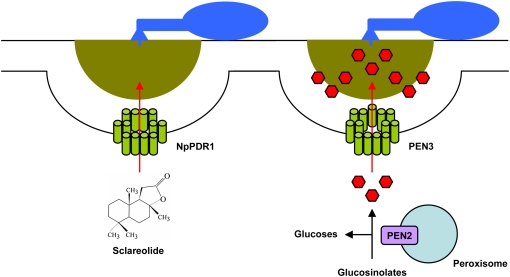
Pathogen-triggered and ABC transporter-driven efflux of small molecules into the apoplast. Plants secret a wide range of secondary metabolites in response to pathogen challenge. In tobacco, the PDR1 ABC transporter is required for the translocation of sclareolide, a toxic phytochemical (left). In Arabidopsis, the PEN3 ABC transporter is required for preinvasive resistance to a broad range of fungal parasites (right). Genetic and biochemical data suggest that peroxisome-associated PEN2 glycosyl hydrolase generates toxic products from glucosinolates that are translocated into the apoplast by PEN3. Due to the self-cytotoxicity of the secreted secondary metabolites, it is likely that their generation and release may occur in direct proximity to microbial contact sites.
Similar articles
-
Transport and secretion in plant-microbe interactions.Curr Opin Plant Biol. 2007 Dec;10(6):573-9. doi: 10.1016/j.pbi.2007.08.002. Epub 2007 Sep 17. Curr Opin Plant Biol. 2007. PMID: 17875397 Review.
-
Recognition of pathogens by plants.Curr Biol. 2000 Jan 13;10(1):R5-7. doi: 10.1016/s0960-9822(99)00273-0. Curr Biol. 2000. PMID: 10660284 Review. No abstract available.
-
Plant science. Auxin begins to give up its secrets.Science. 2006 Sep 1;313(5791):1230-1. doi: 10.1126/science.313.5791.1230. Science. 2006. PMID: 16946049 No abstract available.
-
Cooperative ethylene receptor signaling.Plant Signal Behav. 2012 Aug;7(8):1009-13. doi: 10.4161/psb.20937. Epub 2012 Jul 25. Plant Signal Behav. 2012. PMID: 22827938 Free PMC article. Review.
-
Q&A: How do plants respond to ethylene and what is its importance?BMC Biol. 2016 Jan 27;14:7. doi: 10.1186/s12915-016-0230-0. BMC Biol. 2016. PMID: 26819080 Free PMC article.
Cited by
-
High-throughput confocal imaging of intact live tissue enables quantification of membrane trafficking in Arabidopsis.Plant Physiol. 2010 Nov;154(3):1096-104. doi: 10.1104/pp.110.160325. Epub 2010 Sep 14. Plant Physiol. 2010. PMID: 20841454 Free PMC article.
-
A highway for war and peace: the secretory pathway in plant-microbe interactions.Mol Plant. 2011 Jul;4(4):581-7. doi: 10.1093/mp/ssr053. Epub 2011 Jul 7. Mol Plant. 2011. PMID: 21742620 Free PMC article. Review.
-
A Simple Method for Measuring Apoplast Hydration and Collecting Apoplast Contents.Plant Physiol. 2019 Apr;179(4):1265-1272. doi: 10.1104/pp.18.01076. Epub 2019 Mar 1. Plant Physiol. 2019. PMID: 30824565 Free PMC article.
-
Time-resolved dual transcriptomics reveal early induced Nicotiana benthamiana root genes and conserved infection-promoting Phytophthora palmivora effectors.BMC Biol. 2017 May 11;15(1):39. doi: 10.1186/s12915-017-0379-1. BMC Biol. 2017. PMID: 28494759 Free PMC article.
-
Induction of DREB2A pathway with repression of E2F, jasmonic acid biosynthetic and photosynthesis pathways in cold acclimation-specific freeze-resistant wheat crown.Funct Integr Genomics. 2013 Mar;13(1):57-65. doi: 10.1007/s10142-012-0303-2. Epub 2012 Dec 20. Funct Integr Genomics. 2013. PMID: 23262780
References
-
- Aist JR (1976) Papillae and related wound plugs of plant cells. Annu Rev Phytopathol 14 145–163
-
- Bais HP, Prithiviraj B, Jha AK, Ausubel FM, Vivanco JM (2005) Mediation of pathogen resistance by exudation of antimicrobials from roots. Nature 434 217–221 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
