

Neural and molecular dissection of a C. elegans sensory circuit that regulates fat and feeding
- PMID: 18680713
- PMCID: PMC2556218
- DOI: 10.1016/j.cmet.2008.06.005
Neural and molecular dissection of a C. elegans sensory circuit that regulates fat and feeding
Abstract
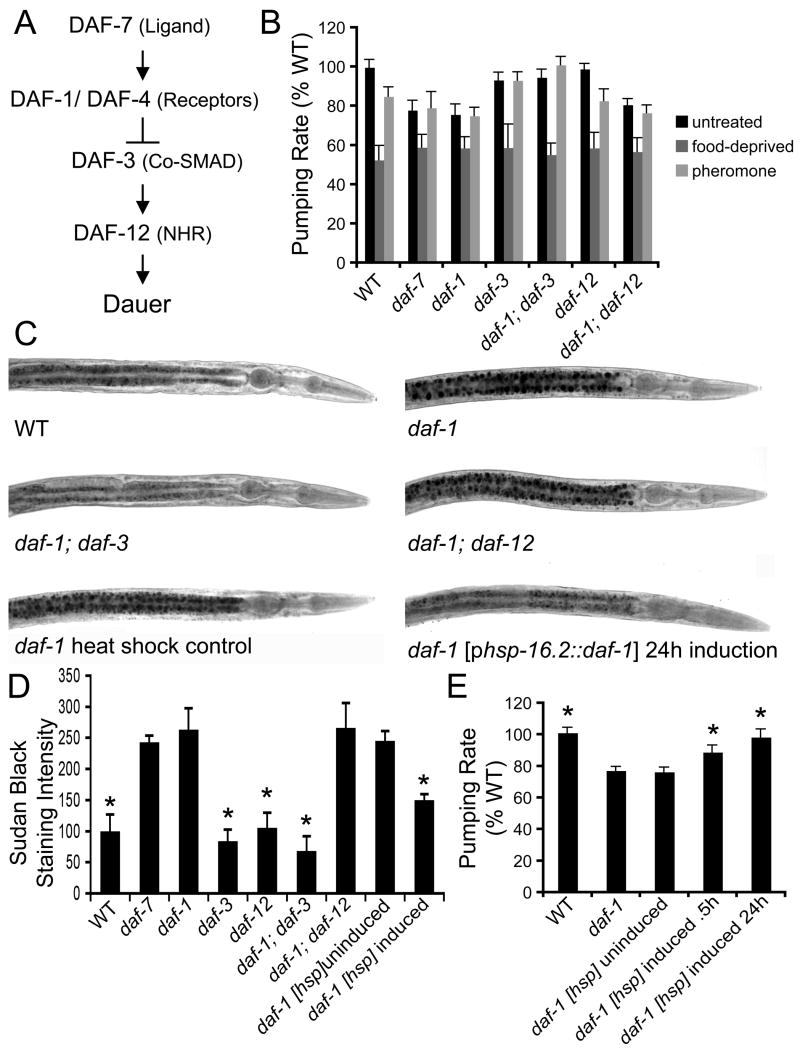
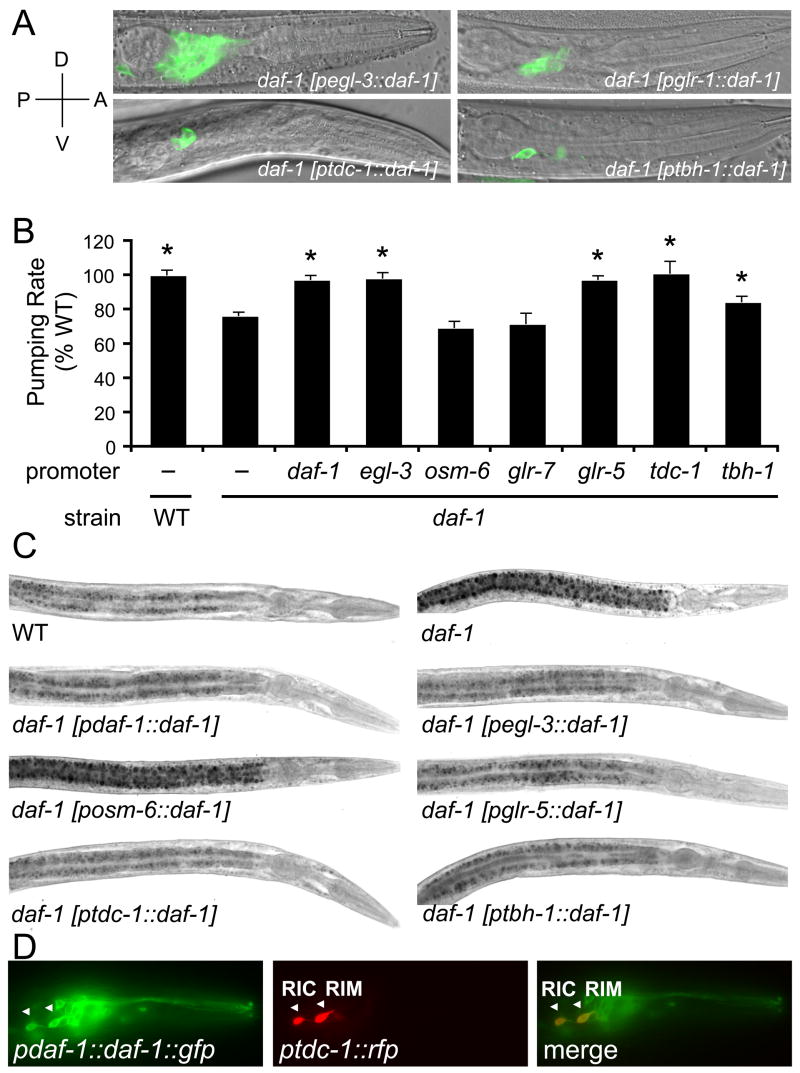
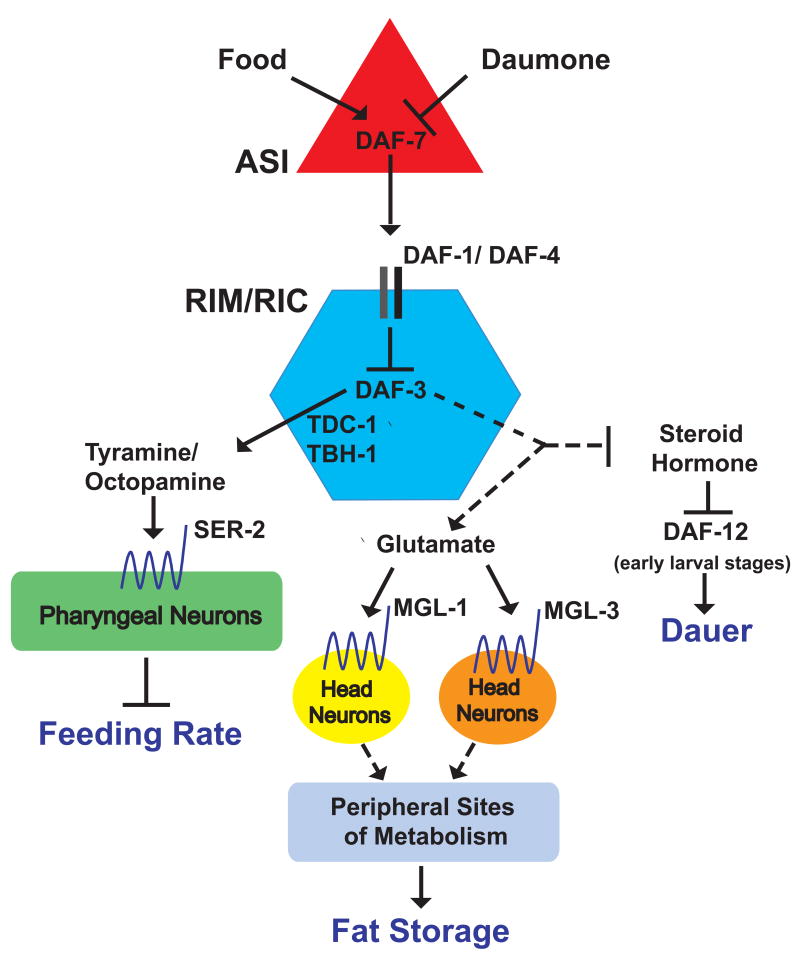
A major challenge in understanding energy balance is deciphering the neural and molecular circuits that govern behavioral, physiological, and metabolic responses of animals to fluctuating environmental conditions. The neurally expressed TGF-beta ligand DAF-7 functions as a gauge of environmental conditions to modulate energy balance in C. elegans. We show that daf-7 signaling regulates fat metabolism and feeding behavior through a compact neural circuit that allows for integration of multiple inputs and the flexibility for differential regulation of outputs. In daf-7 mutants, perception of depleting food resources causes fat accumulation despite reduced feeding rate. This fat accumulation is mediated, in part, through neural metabotropic glutamate signaling and upregulation of peripheral endogenous biosynthetic pathways that direct energetic resources into fat reservoirs. Thus, neural perception of adverse environmental conditions can promote fat accumulation without a concomitant increase in feeding rate.
Figures



Comment in
-
Fattening up without overeating.Cell Metab. 2008 Aug;8(2):95-6. doi: 10.1016/j.cmet.2008.07.003. Cell Metab. 2008. PMID: 18680709 Free PMC article.
References
-
- Albert PS, Riddle DL. Mutants of Caenorhabditis elegans that form dauer-like larvae. Dev Biol. 1988;126:270–293. - PubMed
-
- Alfonso A, Grundahl K, Duerr JS, Han HP, Rand JB. The Caenorhabditis elegans unc-17 gene: a putative vesicular acetylcholine transporter. Science. 1993;261:617–619. - PubMed
-
- Alkema MJ, Hunter-Ensor M, Ringstad N, Horvitz HR. Tyramine Functions independently of octopamine in the Caenorhabditis elegans nervous system. Neuron. 2005;46:247–260. - PubMed
-
- Ansley SJ, Badano JL, Blacque OE, Hill J, Hoskins BE, Leitch CC, Kim JC, Ross AJ, Eichers ER, Teslovich TM, Mah AK, Johnsen RC, Cavender JC, Lewis RA, Leroux MR, Beales PL, Katsanis N. Basal body dysfunction is a likely cause of pleiotropic Bardet-Biedl syndrome. Nature. 2003;425:628–633. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
