

RloC: a wobble nucleotide-excising and zinc-responsive bacterial tRNase
- PMID: 18681940
- PMCID: PMC2610378
- DOI: 10.1111/j.1365-2958.2008.06387.x
RloC: a wobble nucleotide-excising and zinc-responsive bacterial tRNase
Abstract
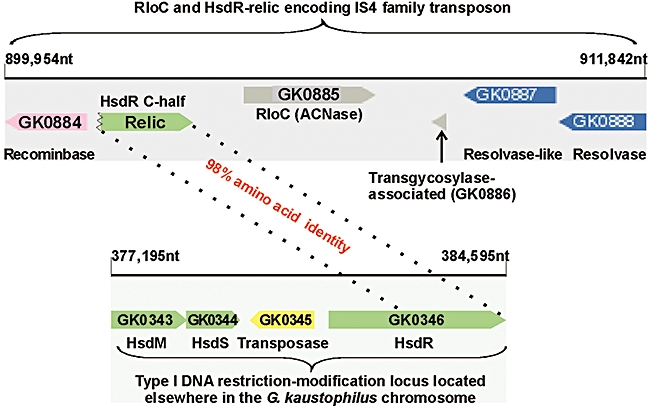
The conserved bacterial protein RloC, a distant homologue of the tRNA(Lys) anticodon nuclease (ACNase) PrrC, is shown here to act as a wobble nucleotide-excising and Zn(++)-responsive tRNase. The more familiar PrrC is silenced by a genetically linked type I DNA restriction-modification (R-M) enzyme, activated by a phage anti-DNA restriction factor and counteracted by phage tRNA repair enzymes. RloC shares PrrC's ABC ATPase motifs and catalytic ACNase triad but features a distinct zinc-hook/coiled-coil insert that renders its ATPase domain similar to Rad50 and related DNA repair proteins. Geobacillus kaustophilus RloC expressed in Escherichia coli exhibited ACNase activity that differed from PrrC's in substrate preference and ability to excise the wobble nucleotide. The latter specificity could impede reversal by phage tRNA repair enzymes and account perhaps for RloC's more frequent occurrence. Mutagenesis and functional assays confirmed RloC's catalytic triad assignment and implicated its zinc hook in regulating the ACNase function. Unlike PrrC, RloC is rarely linked to a type I R-M system but other genomic attributes suggest their possible interaction in trans. As DNA damage alleviates type I DNA restriction, we further propose that these related perturbations prompt RloC to disable translation and thus ward off phage escaping DNA restriction during the recovery from DNA damage.
Figures
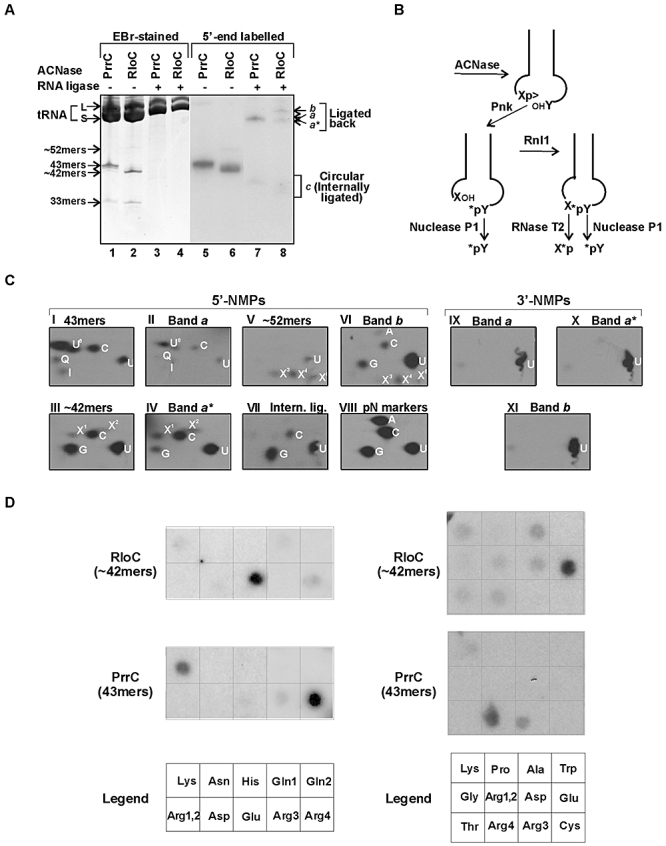

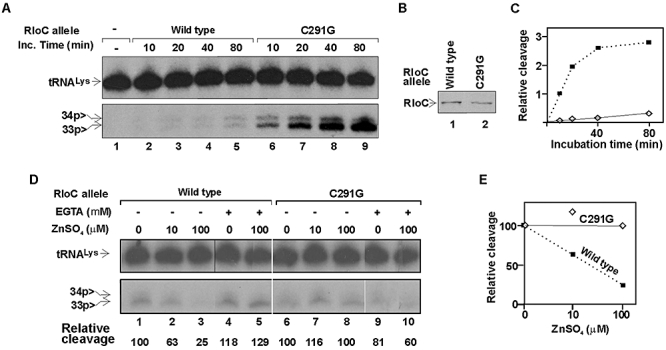
 and second anticodon base U35 indicated. RloC cleaves the substrate initially 5′ to U35 yielding a 5′-fragment containing the radiolabel at the internal 33p34 position. The second cleavage by RloC at PrrC's site exposes the label to the phosphodiesterase/monoesterase activities of T4 Pnk (CPD) but can be pre-empted by prior 3′-dephosphorylation of 34p>. Nuclease P1 releases the indicated end groups from the various labelled products generated by RloC with or without CPD.
and second anticodon base U35 indicated. RloC cleaves the substrate initially 5′ to U35 yielding a 5′-fragment containing the radiolabel at the internal 33p34 position. The second cleavage by RloC at PrrC's site exposes the label to the phosphodiesterase/monoesterase activities of T4 Pnk (CPD) but can be pre-empted by prior 3′-dephosphorylation of 34p>. Nuclease P1 releases the indicated end groups from the various labelled products generated by RloC with or without CPD.
References
-
- Ambudkar SV, Kim IW, Xia D, Sauna ZE. The A-loop, a novel conserved aromatic acid subdomain upstream of the Walker A motif in ABC transporters, is critical for ATP binding. FEBS Lett. 2006;580:1049–1055. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
