

Loss of desmocollin 3 in mice leads to epidermal blistering
- PMID: 18682494
- PMCID: PMC2659849
- DOI: 10.1242/jcs.031518
Loss of desmocollin 3 in mice leads to epidermal blistering
Abstract
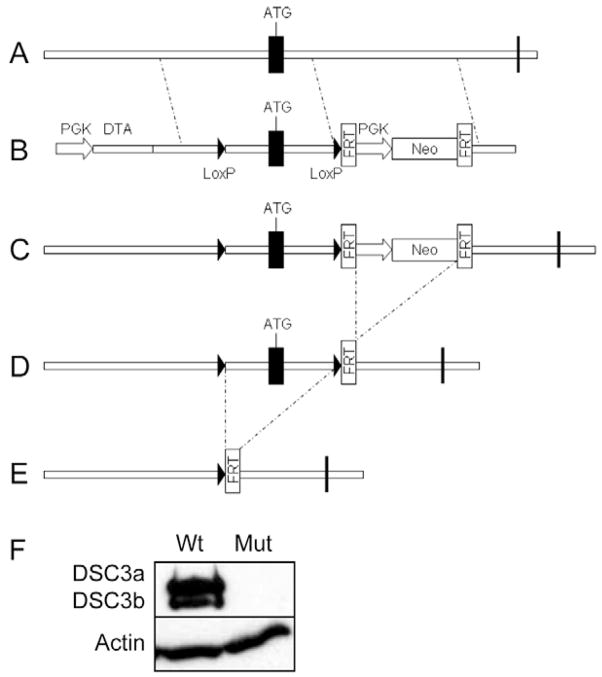
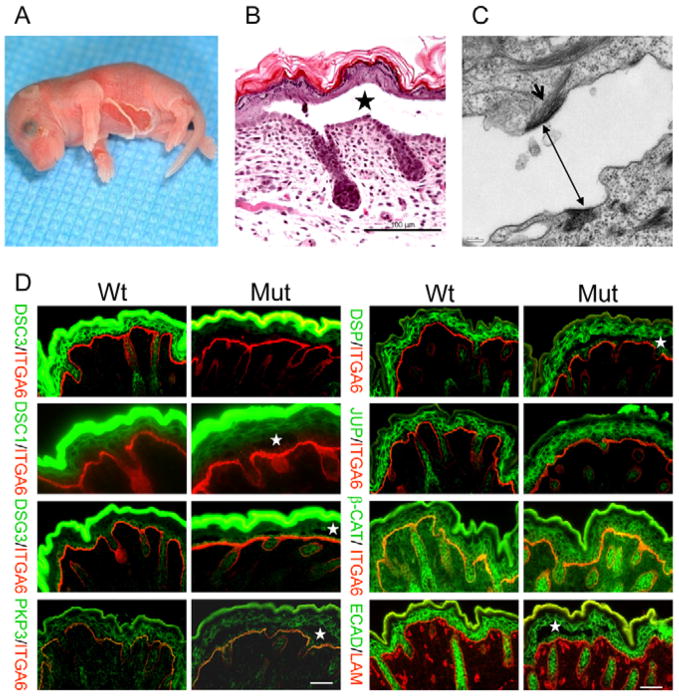
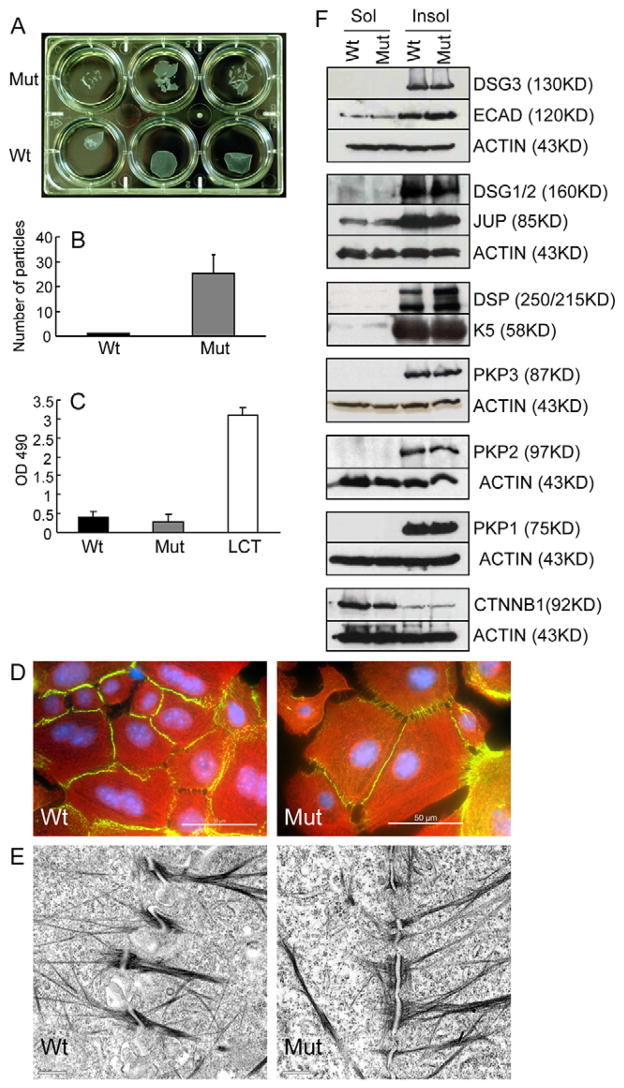
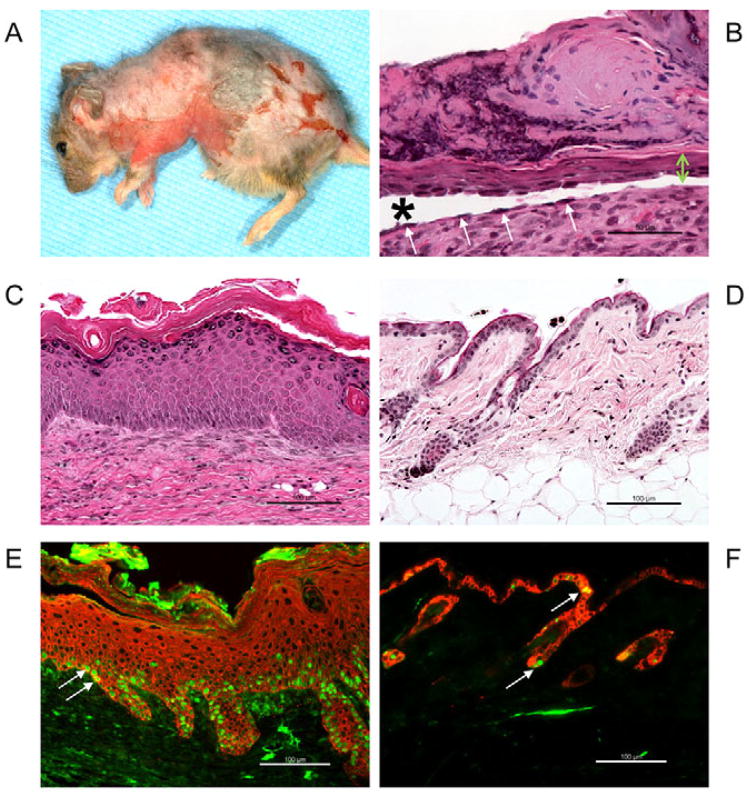
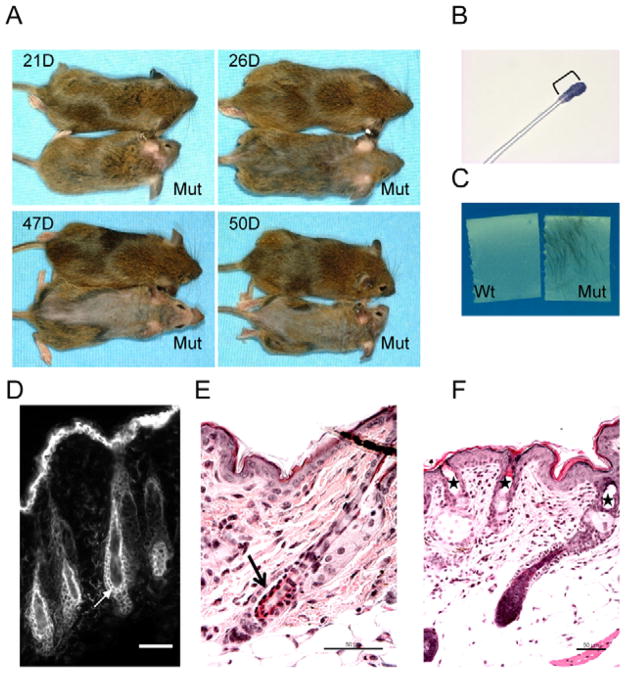
Desmocollin 3 (DSC3) belongs to a subfamily of cadherins and is a major component of desmosomes in keratinocytes of stratified epithelia, such as the epidermis. Based on its amino acid sequence homology to classical cadherins, such as E-cadherin, it has been postulated that DSC3 functions as a cell-adhesion molecule. To test this hypothesis, we assessed the function of DSC3 in the development and maintenance of stratified epithelia, in particular the epidermis and hair follicles. Using a conditional null allele, we show that loss of Dsc3 function in the epidermis causes impaired cell-cell adhesion, leading to intra-epidermal blistering and telogen hair loss. Furthermore, the lesions in Dsc3-null skin resemble those observed in individuals with pemphigus vulgaris (PV), indicating that impaired Dsc3 function could be a potential cause of PV-like inherited or acquired skin blistering diseases.
Figures
References
-
- Amagai M, Ahmed AR, Kitajima Y, Bystryn JC, Milner Y, Gniadecki R, Hertl M, Pincelli C, Kurzen H, Fridkis-Hareli M, et al. Are desmoglein autoantibodies essential for the immunopathogenesis of pemphigus vulgaris, or just “witnesses of disease”? Exp Dermatol. 2006;15:815–831. - PubMed
-
- Bolling MC, Mekkes JR, Goldschmidt WF, van Noesel CJ, Jonkman MF, Pas HH. Acquired palmoplantar keratoderma and immunobullous disease associated with antibodies to desmocollin 3. Br J Dermatol. 2007;157:168–173. - PubMed
-
- Chen J, Cheng X, Merched-Sauvage M, Caulin C, Roop DR, Koch PJ. An unexpected role for keratin 10 end domains in susceptibility to skin cancer. J Cell Sci. 2006;119:5067–5076. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
