

Nonhuman primates as models for human adrenal androgen production: function and dysfunction
- PMID: 18683055
- PMCID: PMC2653599
- DOI: 10.1007/s11154-008-9099-8
Nonhuman primates as models for human adrenal androgen production: function and dysfunction
Abstract
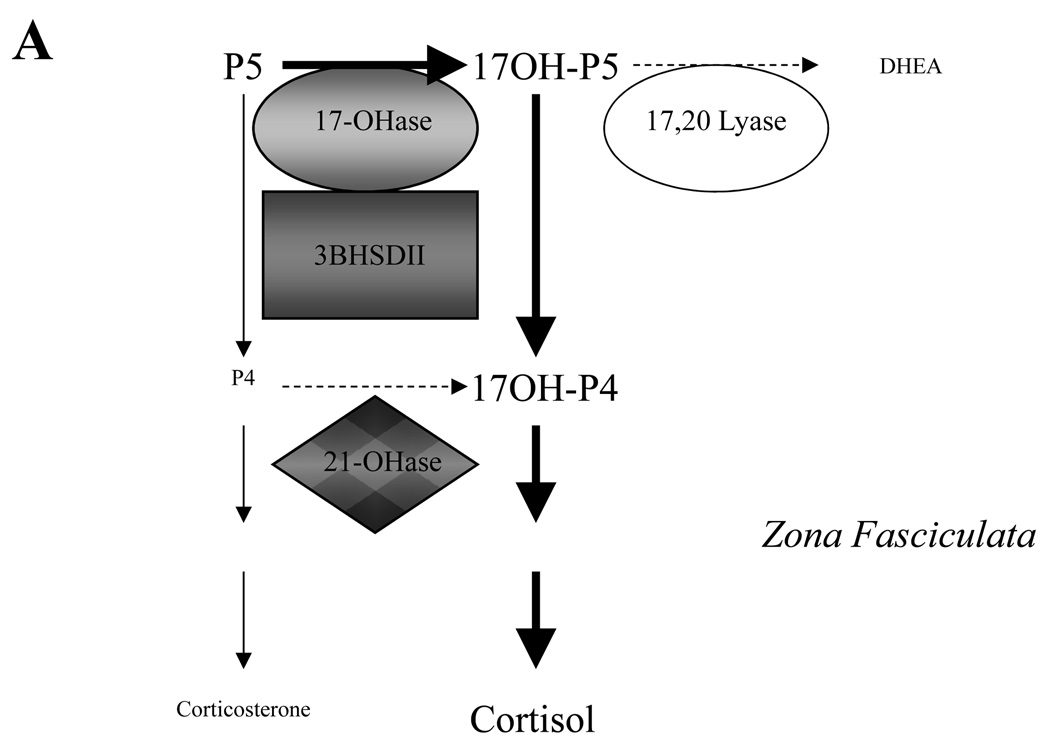
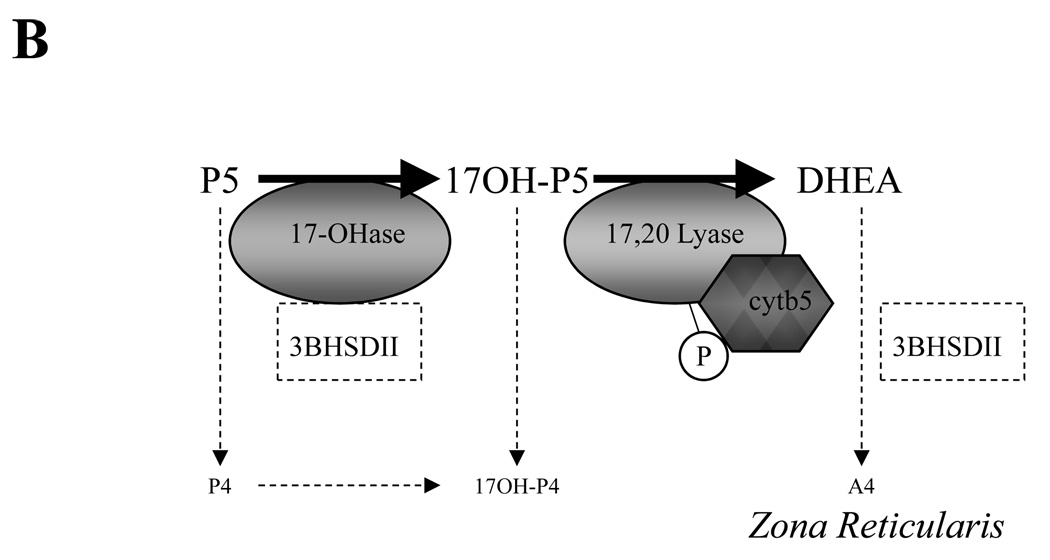
The origin of circulating DHEA and adrenal-derived androgens in humans and nonhuman primates is largely distinct from other mammalian species. In humans and many Old world primates, the fetal adrenal gland and adult zona reticularis (ZR) are known to be the source for production of DHEA (and DHEAS) in mg quantities. In spite of similarities there are also some differences. Herein, we take a comparative endocrine approach to the diversity of adrenal androgen biosynthesis and its developmental timing in three primate species to illustrate how understanding such differences may provide unique insight into mechanisms underlying adrenal androgen regulation and its pathophysiology in humans. We contrast the conventional developmental onset of adrenal DHEA biosynthesis at adrenarche in humans with (1) an earlier, peri-partutrition onset of adrenal DHEA synthesis in rhesus macaques (Old World primate) and (2) a more dynamic and reversible onset of adrenal DHEA biosynthesis in female marmosets (New World primate), and further consider these events in terms of the corresponding developmental changes in expression of CYP17, HSD3B2 and CYB5 in the ZR. We also integrate these observations with recently described biochemical characterization of CYP17 cDNA cloned from each of these nonhuman primate species and the corresponding effects of phosphorylation versus CYB5 coexpression on 17,20 lyase versus 17-hydroxylase activity in each case. In addition, female rhesus macaques exposed in utero to exogenous androgen excess, exhibit symptoms of adrenal hyperandrogenism in adult females in a manner reminiscent of that seen in the human condition of PCOS. The possible mechanisms underlying such adrenal hyperandrogenism are further considered in terms of the effects of altered relative expression of CYP17, HSD3B2 and CYB5 as well as the altered signaling responses of various kinases including protein kinase A, or the insulin sensitive PI3-kinase/AKT signaling pathway which may impact on 17,20 lyase activity. We conclude that while the triggers for the onset of ZR function in all three species show clear differences (age, stage of development, social status, gender), there are still common mechanisms driving an increase in DHEA biosynthesis in each case. A full understanding of the mechanisms that control 17,20 lyase function and dysfunction in humans may best be achieved by comparative studies of the endocrine mechanisms controlling adrenal ZR function and dysfunction in these nonhuman primate species.
Figures
References
-
- Abbott DH, Barnett DK, Bruns CM, Dumesic DA. Androgen excess fetal programming of female reproduction: a developmental aetiology for polycystic ovary syndrome? Hum Reprod Update. 2005;11:357–374. - PubMed
-
- Abbott DH, Barnett DK, Bruns CM, Dunaif A, Dumesic DA. Transient hyperglycemia in both mother and fetus from experimental induction of maternal androgen excess in a nonhuman primate model for polycystic ovary syndrome (abstract P3-113). 88th Annual Meeting of the Endocrine Society, Boston; 2006.
-
- Abbott DH, Dumesic DA, Levine JE, Dunaif A, Padmanabhan V. Animal models and fetal programming of PCOS. In: Azziz R, Nestler JE, Dewailly D, editors. Contemporary Endocrinology: Androgen Excess Disorders in Women: Polycystic Ovary Syndrome and Other Disorders. 2nd Edition. Totowa, NJ: Humana Press Inc.; 2006. pp. 259–272.
-
- Abbott DH, Goodfriend TL, Dunaid A, Muller SJ, Dumesic DA, Tarantal AF. Increased body weight and enhanced insulin sensitivity in infant female rhesus monkeys exposed to androgen excess during early gestation; Abstract P2-348 presented at the 89th Annual Meeting of the Endocrine Society, Toronto, Canada, June 2–5; 2007.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
