

ICP34.5-dependent and -independent activities of salubrinal in herpes simplex virus-1 infected cells
- PMID: 18684481
- PMCID: PMC2665023
- DOI: 10.1016/j.virol.2008.06.028
ICP34.5-dependent and -independent activities of salubrinal in herpes simplex virus-1 infected cells
Abstract
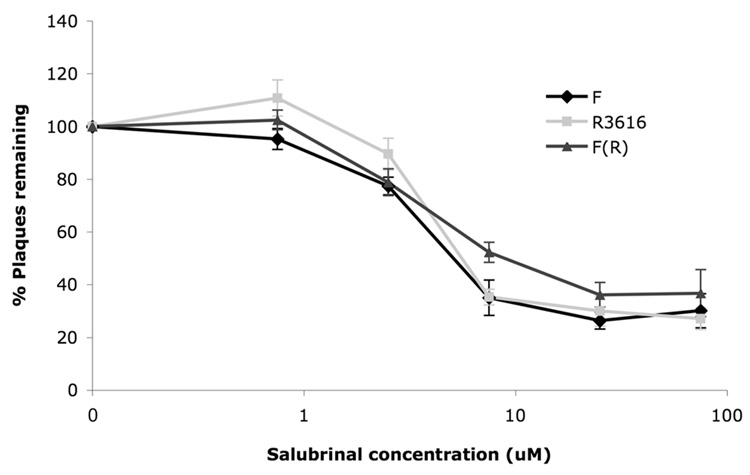
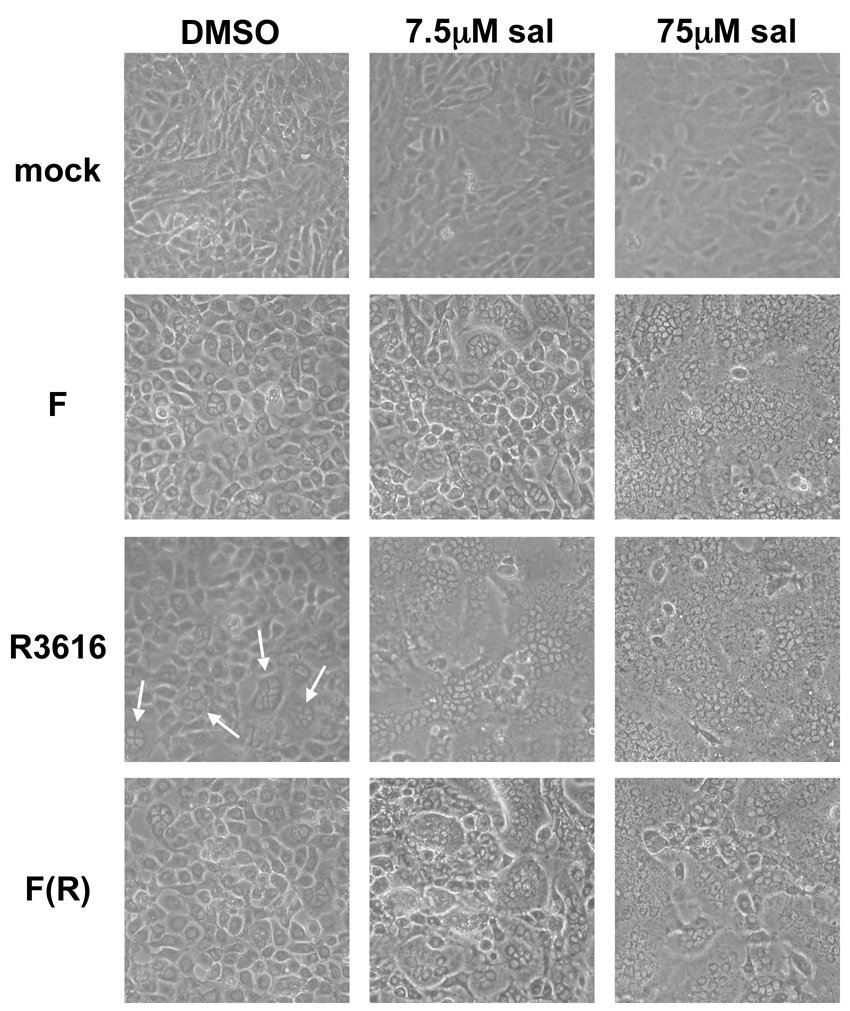
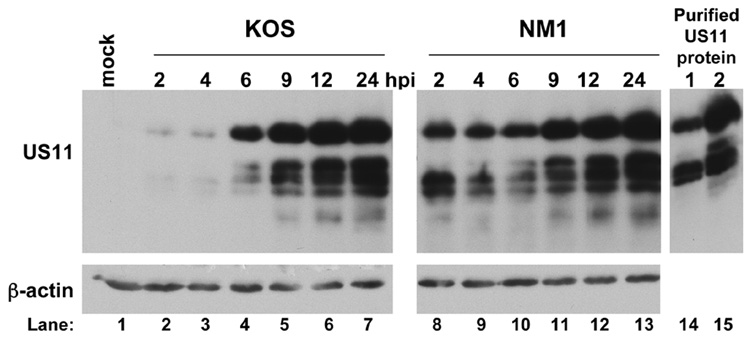
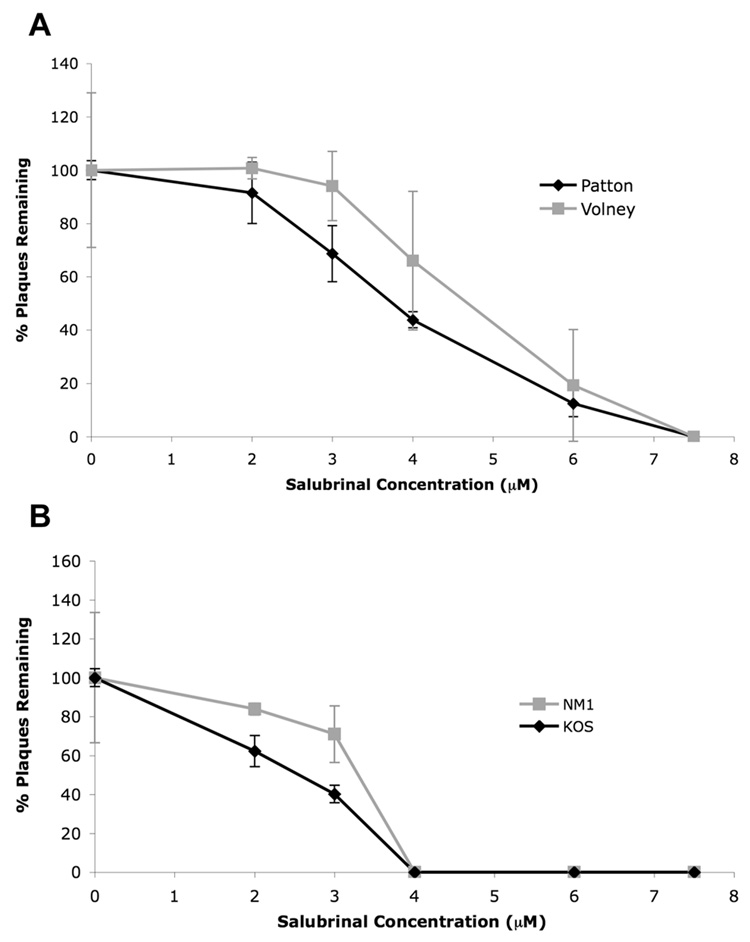
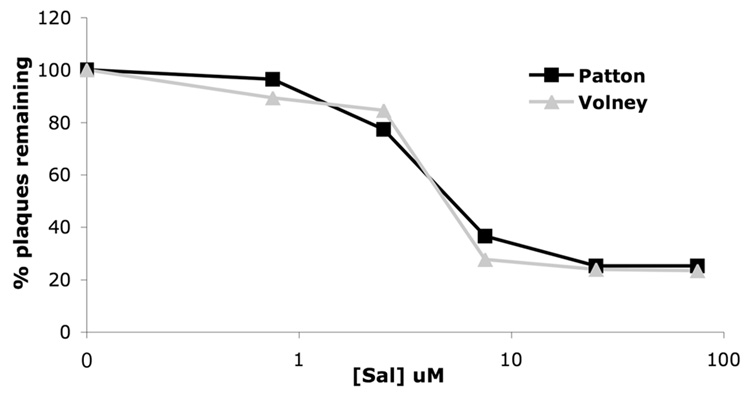
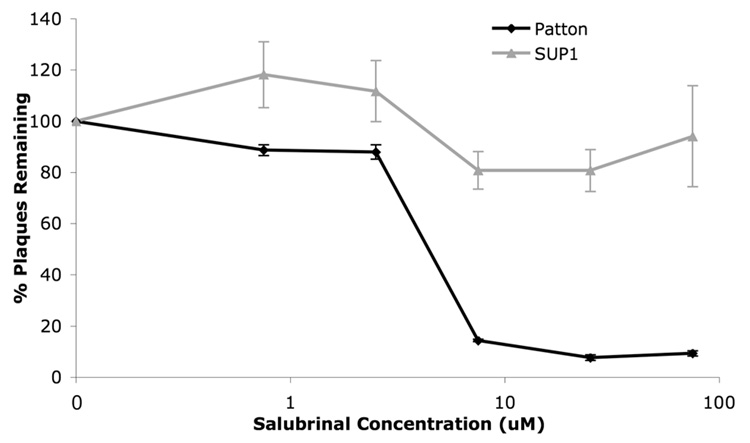
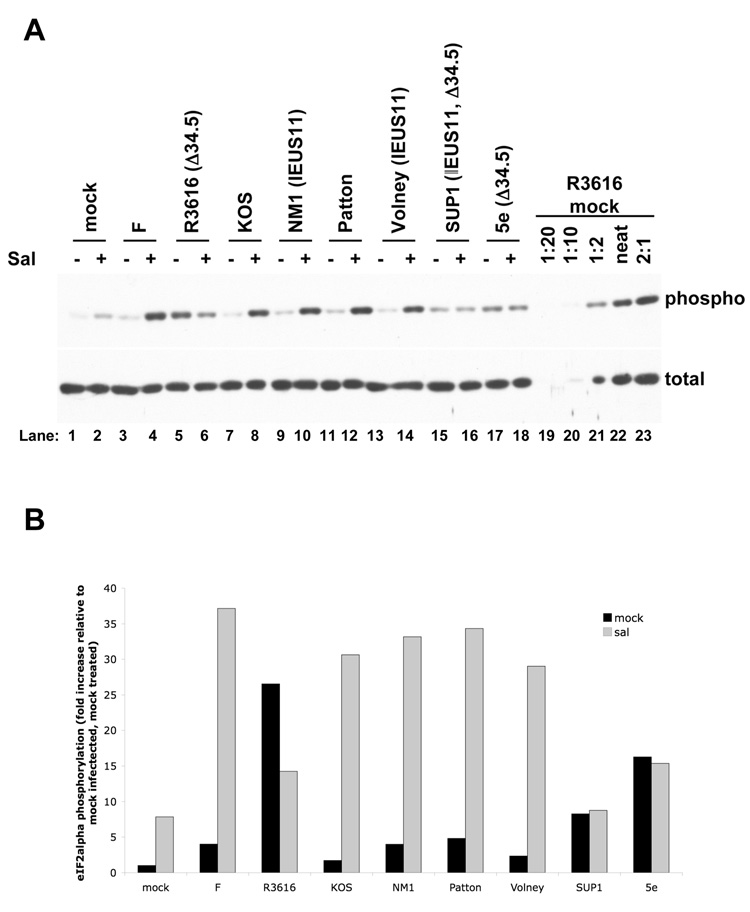
The small molecule salubrinal has antiviral activity against herpes simplex virus-1 (HSV-1) and inhibits dephosphorylation of eIF2 alpha mediated by the HSV-1 protein ICP34.5. We investigated whether salubrinal's activities in infected cells depend on ICP34.5. An ICP34.5 deletion mutant was as sensitive as wild type HSV-1 to salubrinal inhibition of plaque formation in Vero cells. However, salubrinal induced formation of syncytia in infected Vero cells, which was enhanced by ICP34.5 mutations. Expression of HSV-1 US11 with immediate early kinetics, which is known to suppress the effects of ICP34.5 mutations, resulted in slight resistance to salubrinal in murine embryonic fibroblasts, and substantial resistance in those cells when ICP34.5 was additionally mutated. ICP34.5 mutations, but not immediate early expression of US11, prevented salubrinal's ability to increase phosphorylation of eIF2 alpha during HSV-1 infection of Vero cells. Taken together, our data indicate that salubrinal has both ICP34.5-dependent and -independent activities in HSV-1 infected cells.
Figures
References
-
- Boyce M, Bryant KF, Jousse C, Long K, Harding HP, Scheuner D, Kaufman RJ, Ma D, Coen DM, Ron D, Yuan J. A selective inhibitor of eIF2alpha dephosphorylation protects cells from ER stress. Science. 2005;307:935–939. - PubMed
-
- Boyce M, Yuan J. Cellular response to endoplasmic reticulum stress: a matter of life or death. Cell Death Differ. 2006;13:363–373. - PubMed
-
- Brown SM, MacLean AR, Aitken JD, Harland J. ICP34.5 influences herpes simplex virus type 1 maturation and egress from infected cells in vitro. J Gen Virol. 1994;75:3679–3686. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
