

Retinoic acid increases Foxp3+ regulatory T cells and inhibits development of Th17 cells by enhancing TGF-beta-driven Smad3 signaling and inhibiting IL-6 and IL-23 receptor expression
- PMID: 18684916
- PMCID: PMC2722959
- DOI: 10.4049/jimmunol.181.4.2277
Retinoic acid increases Foxp3+ regulatory T cells and inhibits development of Th17 cells by enhancing TGF-beta-driven Smad3 signaling and inhibiting IL-6 and IL-23 receptor expression
Abstract
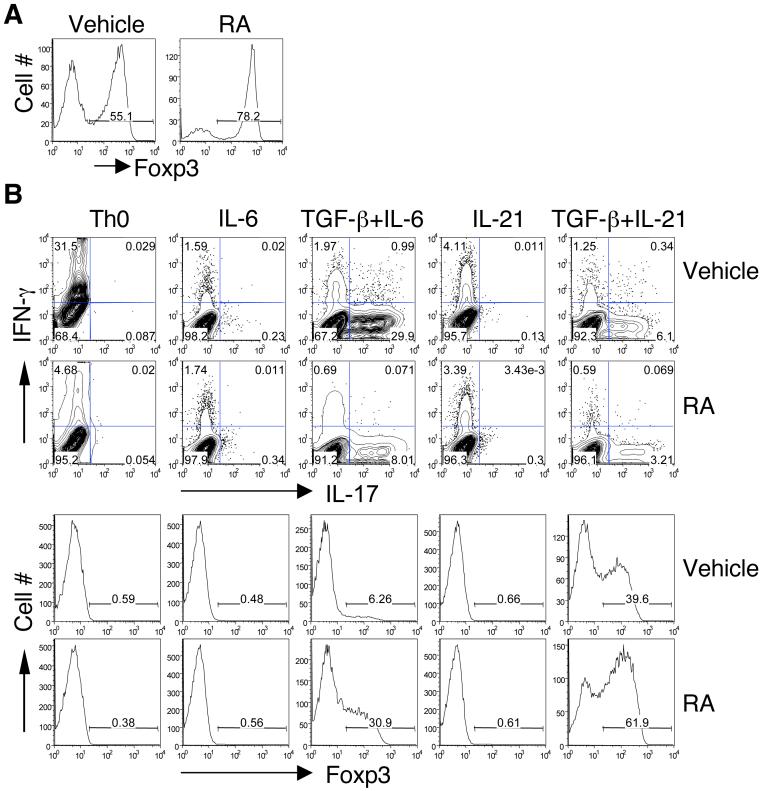
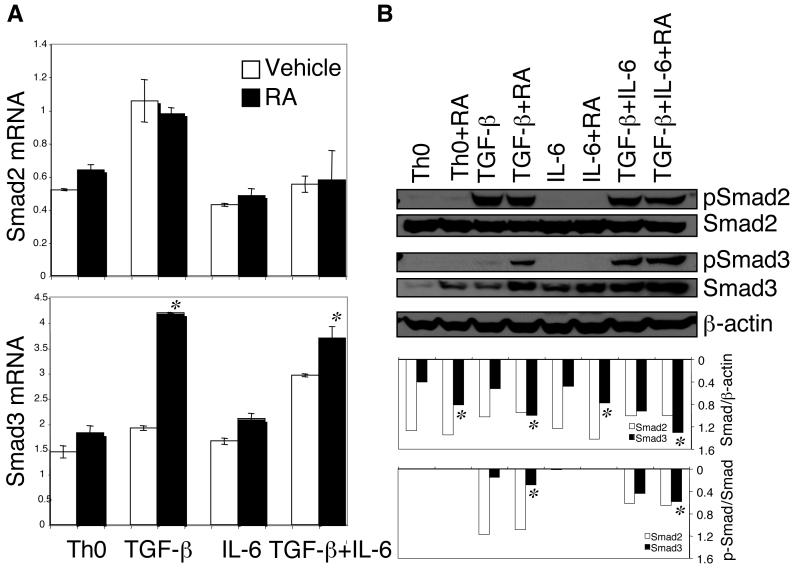
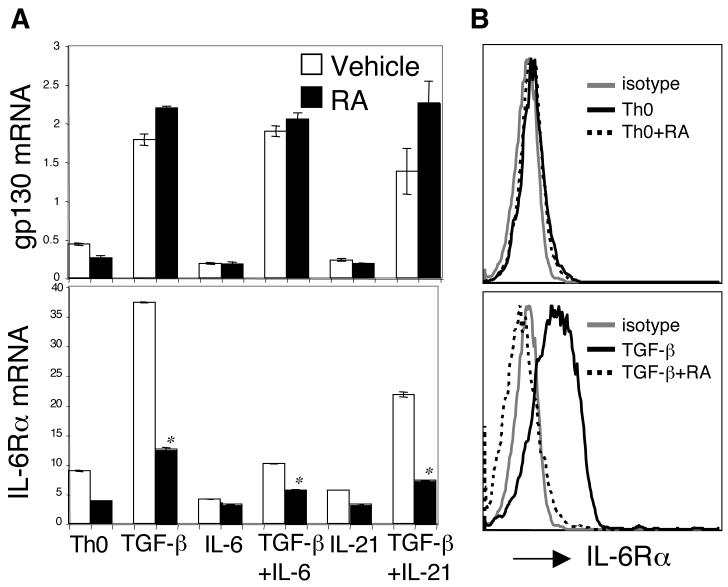
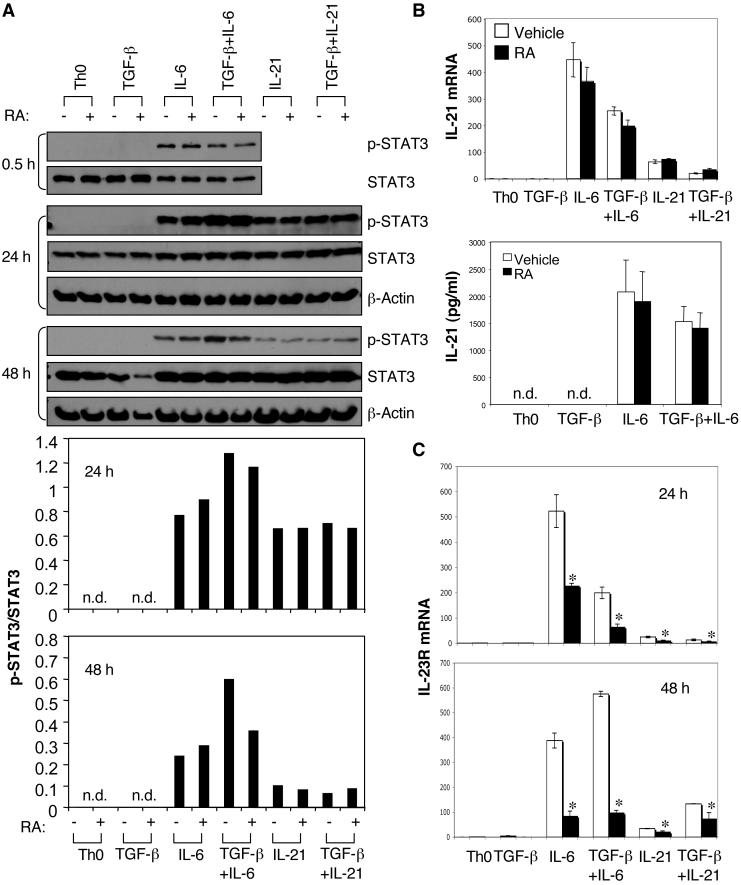
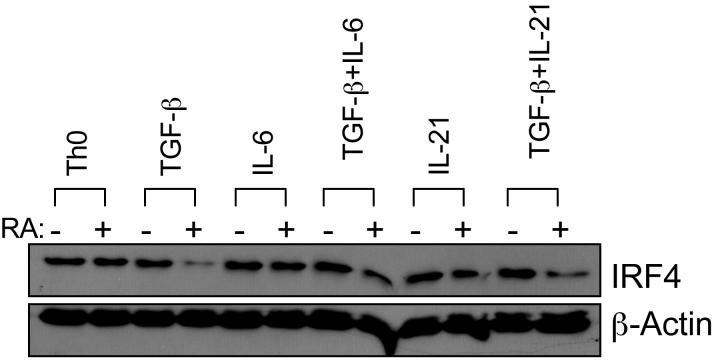
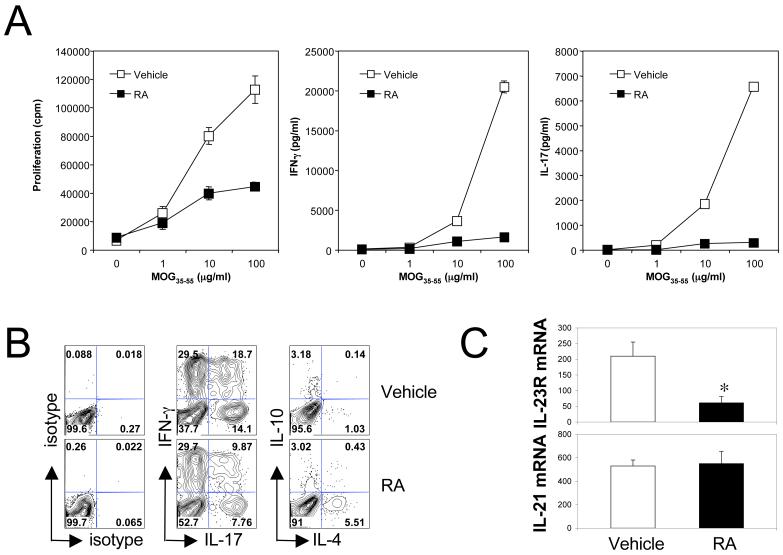
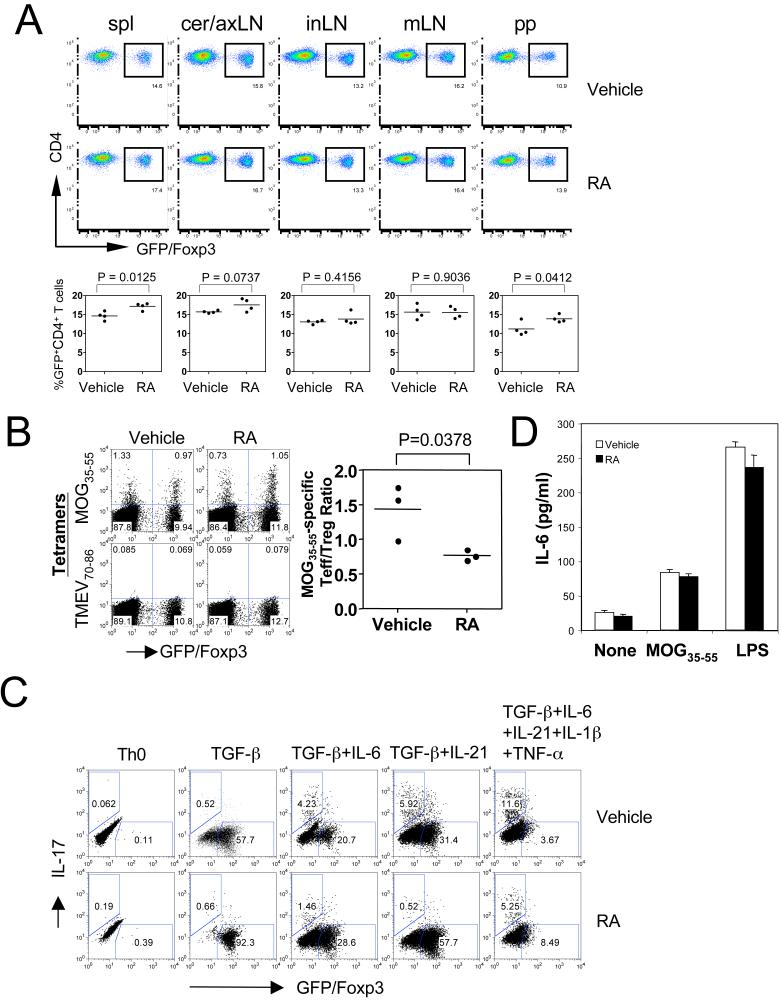
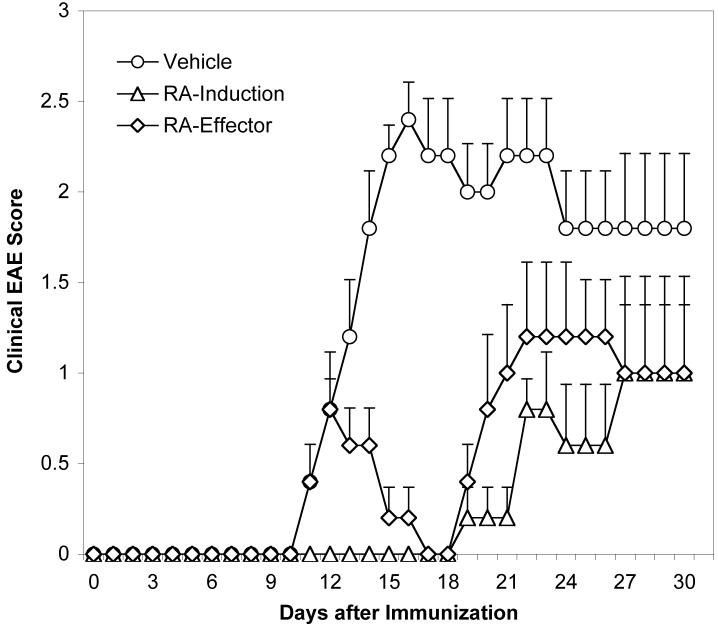
The de novo generation of Foxp3+ regulatory T (Treg) cells in the peripheral immune compartment and the differentiation of Th17 cells both require TGF-beta, and IL-6 and IL-21 are switch factors that drive the development of Th17 cells at the expense of Treg cell generation. The major vitamin A metabolite all-trans retinoic acid (RA) not only enforces the generation of Treg cells but also inhibits the differentiation of Th17 cells. Herein we show that RA enhances TGF-beta signaling by increasing the expression and phosphorylation of Smad3, and this results in increased Foxp3 expression even in the presence of IL-6 or IL-21. RA also inhibits the expression of IL-6Ralpha, IRF-4, and IL-23R and thus inhibits Th17 development. In vitro, RA significantly promotes Treg cell conversion, but in vivo during the development of experimental autoimmune encephalomyelitis it does not increase the frequency of Treg cells in the face of an ongoing inflammation. However, RA suppresses the disease very efficiently by inhibiting proinflammatory T cell responses, especially pathogenic Th17 responses. These data not only identify the signaling mechanisms by which RA can affect both Treg cell and Th17 differentiation, but they also highlight that in vivo during an autoimmune reaction, RA suppresses autoimmunity mainly by inhibiting the generation of effector Th17 cells.
Figures
References
-
- Abbas AK, Murphy KM, Sher A. Functional diversity of helper T lymphocytes. Nature. 1996;383:787. - PubMed
-
- Harrington LE, Mangan PR, Weaver CT. Expanding the effector CD4 T-cell repertoire: the Th17 lineage. Curr Opin Immunol. 2006;18:349. - PubMed
-
- Bettelli E, Oukka M, Kuchroo VK. T(H)-17 cells in the circle of immunity and autoimmunity. Nat Immunol. 2007;8:345. - PubMed
-
- Korn T, Oukka M. Dynamics of antigen-specific regulatory T-cells in the context of autoimmunity. Semin Immunol. 2007;19:272. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
