

Deficiency in ubiquitin ligase TRIM2 causes accumulation of neurofilament light chain and neurodegeneration
- PMID: 18687884
- PMCID: PMC2575299
- DOI: 10.1073/pnas.0802261105
Deficiency in ubiquitin ligase TRIM2 causes accumulation of neurofilament light chain and neurodegeneration
Abstract
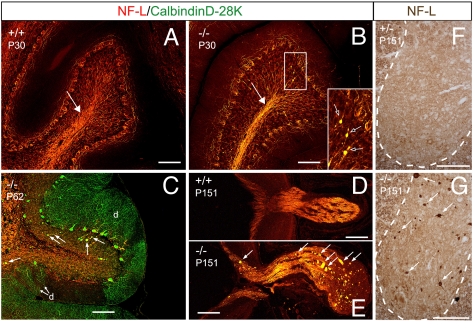
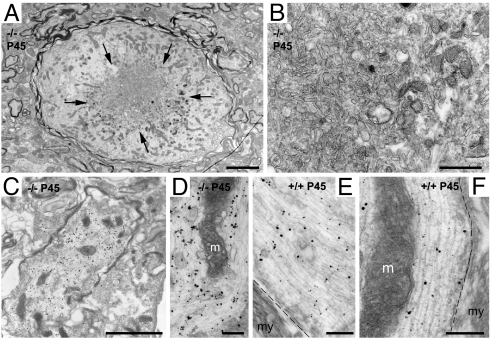
TRIM RING finger proteins have been shown to play an important role in cancerogenesis, in the pathogenesis of some human hereditary disorders, and in the defense against viral infection, but the function of the majority of TRIM proteins remains unknown. Here, we show that TRIM RING finger protein TRIM2, highly expressed in the nervous system, is an UbcH5a-dependent ubiquitin ligase. We further demonstrate that TRIM2 binds to neurofilament light subunit (NF-L) and regulates NF-L ubiquitination. Additionally, we show that mice deficient in TRIM2 have increased NF-L level in axons and NF-L-filled axonal swellings in cerebellum, retina, spinal cord, and cerebral cortex. The axonopathy is followed by progressive neurodegeneration accompanied by juvenile-onset tremor and ataxia. Our results demonstrate that TRIM2 is an ubiquitin ligase and point to a mechanism regulating NF-L metabolism through an ubiquitination pathway that, if deregulated, triggers neurodegeneration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
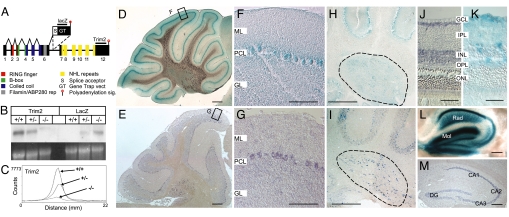
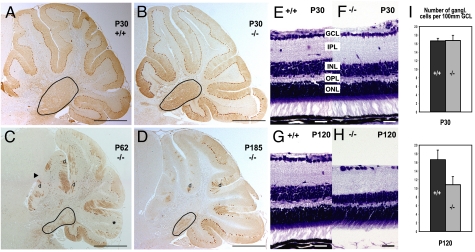
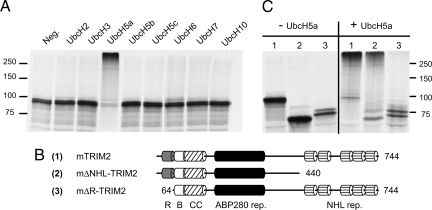
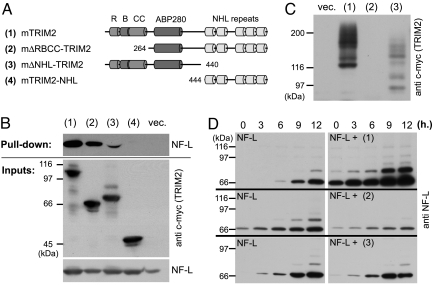
References
-
- Freemont PS. RING for destruction? Curr Biol. 2000;10:R84–R87. - PubMed
-
- de The H, et al. The PML-RAR alpha fusion mRNA generated by the t(15;17) translocation in acute promyelocytic leukemia encodes a functionally altered RAR. Cell. 1991;66:675–684. - PubMed
-
- Kaiser SM, Malik HS, Emerman M. Restriction of an extinct retrovirus by the human TRIM5alpha antiviral protein. Science. 2007;316:1756–1758. - PubMed
-
- Shimura H, et al. Familial Parkinson disease gene product, parkin, is a ubiquitin-protein ligase. Nat Genet. 2000;25:302–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
