

Receptor actions of synaptically released glutamate: the role of transporters on the scale from nanometers to microns
- PMID: 18689452
- PMCID: PMC2576387
- DOI: 10.1529/biophysj.108.129874
Receptor actions of synaptically released glutamate: the role of transporters on the scale from nanometers to microns
Abstract
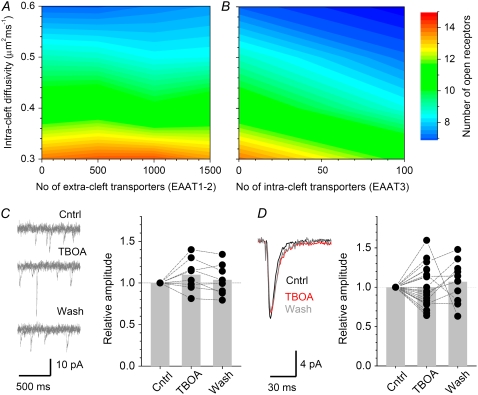
Actions of the excitatory neurotransmitter glutamate inside and outside the synaptic cleft determine the activity of neural circuits in the brain. However, to what degree local glutamate transporters affect these actions on a submicron scale remains poorly understood. Here we focus on hippocampal area CA1, a common subject of synaptic physiology studies. First, we use a two-photon excitation technique to obtain an estimate of the apparent (macroscopic) extracellular diffusion coefficient for glutamate, approximately 0.32 mum(2)/ms. Second, we incorporate this measurement into a Monte Carlo model of the typical excitatory synapse and examine the influence of distributed glutamate transporter molecules on signal transmission. Combined with the results of whole-cell recordings, such simulations argue that, although glutamate transporters have little effect on the activation of synaptic alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors, this does not rule out the occurrence of up to several dozens of transporters inside the cleft. We further evaluate how the expression pattern of transporter molecules (on the 10-100 nm scale) affects the activation of N-methyl-D-aspartic acid or metabotropic glutamate receptors in the synaptic vicinity. Finally, we extend our simulations to the macroscopic scale, estimating that synaptic activity sufficient to excite principal neurons could intermittently raise extracellular glutamate to approximately 1 muM only at sparse (microns apart) hotspots. Greater rises of glutamate occur only when <5% of transporters are available (for instance, when an astrocyte fails). The results provide a quantitative framework for a better understanding of the relationship between glutamate transporters and glutamate receptor signaling.
Figures





Similar articles
-
Synaptically released glutamate does not overwhelm transporters on hippocampal astrocytes during high-frequency stimulation.J Neurophysiol. 2000 May;83(5):2835-43. doi: 10.1152/jn.2000.83.5.2835. J Neurophysiol. 2000. PMID: 10805681
-
Astrocyte glutamate transporters regulate metabotropic glutamate receptor-mediated excitation of hippocampal interneurons.J Neurosci. 2004 May 12;24(19):4551-9. doi: 10.1523/JNEUROSCI.5217-03.2004. J Neurosci. 2004. PMID: 15140926 Free PMC article.
-
Glutamate transporters bring competition to the synapse.Curr Opin Neurobiol. 2004 Jun;14(3):346-52. doi: 10.1016/j.conb.2004.05.007. Curr Opin Neurobiol. 2004. PMID: 15194115 Review.
-
Simulation of postsynaptic glutamate receptors reveals critical features of glutamatergic transmission.PLoS One. 2011;6(12):e28380. doi: 10.1371/journal.pone.0028380. Epub 2011 Dec 15. PLoS One. 2011. PMID: 22194830 Free PMC article.
-
Glutamate transporters: confining runaway excitation by shaping synaptic transmission.Nat Rev Neurosci. 2007 Dec;8(12):935-47. doi: 10.1038/nrn2274. Nat Rev Neurosci. 2007. PMID: 17987031 Review.
Cited by
-
A computational model to investigate astrocytic glutamate uptake influence on synaptic transmission and neuronal spiking.Front Comput Neurosci. 2012 Oct 1;6:70. doi: 10.3389/fncom.2012.00070. eCollection 2012. Front Comput Neurosci. 2012. PMID: 23060782 Free PMC article.
-
Synaptic GABA release prevents GABA transporter type-1 reversal during excessive network activity.Nat Commun. 2015 Mar 23;6:6597. doi: 10.1038/ncomms7597. Nat Commun. 2015. PMID: 25798861 Free PMC article.
-
Disentangling astroglial physiology with a realistic cell model in silico.Nat Commun. 2018 Sep 3;9(1):3554. doi: 10.1038/s41467-018-05896-w. Nat Commun. 2018. PMID: 30177844 Free PMC article.
-
Human neutrophils communicate remotely via calcium-dependent glutamate-induced glutamate release.iScience. 2023 Jun 28;26(7):107236. doi: 10.1016/j.isci.2023.107236. eCollection 2023 Jul 21. iScience. 2023. PMID: 37496680 Free PMC article.
-
Monitoring cell membrane recycling dynamics of proteins using whole-cell fluorescence recovery after photobleaching of pH-sensitive genetic tags.Nat Protoc. 2022 Dec;17(12):3056-3079. doi: 10.1038/s41596-022-00732-4. Epub 2022 Sep 5. Nat Protoc. 2022. PMID: 36064755 Review.
References
-
- Sah, P., S. Hestrin, and R. A. Nicoll. 1989. Tonic activation of NMDA receptros by ambient glutamate enhances excitability of neurons. Science. 246:815–818. - PubMed
-
- Eghbali, M., J. P. Curmi, B. Birnir, and P. W. Gage. 1997. Hippocampal GABA(A) channel conductance increased by diazepam. Nature. 388:71–75. - PubMed
-
- Mitchell, S. J., and R. A. Silver. 2003. Shunting inhibition modulates neuronal gain during synaptic excitation. Neuron. 38:433–435. - PubMed
-
- Semyanov, A., M. C. Walker, D. M. Kullmann, and R. A. Silver. 2004. Tonically active GABA A receptors: modulating gain and maintaining the tone. Trends Neurosci. 27:262–269. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
