

Polar localization and compartmentalization of ClpP proteases during growth and sporulation in Bacillus subtilis
- PMID: 18689476
- PMCID: PMC2566194
- DOI: 10.1128/JB.00589-08
Polar localization and compartmentalization of ClpP proteases during growth and sporulation in Bacillus subtilis
Abstract
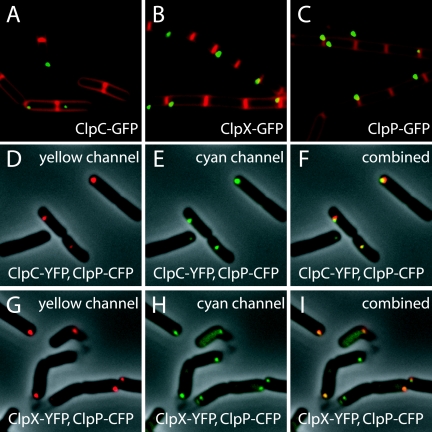
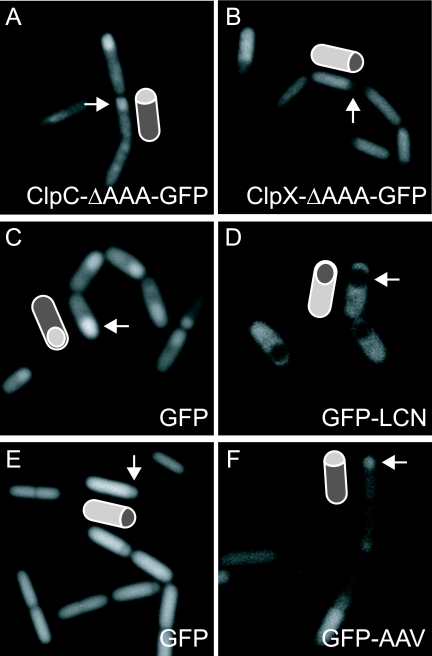
Spatial control of proteolysis is emerging as a common feature of regulatory networks in bacteria. In the spore-forming bacterium Bacillus subtilis, the peptidase ClpP can associate with any of three ATPases: ClpC, ClpE, and ClpX. Here, we report that ClpCP, ClpEP, and ClpXP localize in foci often near the poles of growing cells and that ClpP and the ATPase are each capable of polar localization independently of the other component. A region of ClpC containing an AAA domain was necessary and sufficient for polar localization. We also report that ClpCP and ClpXP proteases differentially localize to the forespore and mother cell compartments of the sporangium during spore formation. Moreover, model substrates for each protease created by appending recognition sequences for ClpCP or ClpXP to the green fluorescent protein were preferentially eliminated from the forespore or the mother cell, respectively. Biased accumulation of ClpCP in the forespore may contribute to the cell-specific activation of the transcription factor sigma(F) by preferential ClpCP-dependent degradation of the anti-sigma(F) factor SpoIIAB.
Figures




Similar articles
-
Clp and Lon proteases occupy distinct subcellular positions in Bacillus subtilis.J Bacteriol. 2008 Oct;190(20):6758-68. doi: 10.1128/JB.00590-08. Epub 2008 Aug 8. J Bacteriol. 2008. PMID: 18689473 Free PMC article.
-
Localization of general and regulatory proteolysis in Bacillus subtilis cells.Mol Microbiol. 2008 Nov;70(3):682-94. doi: 10.1111/j.1365-2958.2008.06438.x. Epub 2008 Sep 10. Mol Microbiol. 2008. PMID: 18786145 Free PMC article.
-
Unique degradation signal for ClpCP in Bacillus subtilis.J Bacteriol. 2003 Sep;185(17):5275-8. doi: 10.1128/JB.185.17.5275-5278.2003. J Bacteriol. 2003. PMID: 12923101 Free PMC article.
-
Chaperone-protease systems in regulation and protein quality control in Bacillus subtilis.Res Microbiol. 2009 Nov;160(9):637-44. doi: 10.1016/j.resmic.2009.08.020. Epub 2009 Sep 23. Res Microbiol. 2009. PMID: 19781636 Review.
-
Compartmentalized transcription and the establishment of cell type during sporulation in Bacillus subtilis.Bioessays. 1992 Feb;14(2):125-8. doi: 10.1002/bies.950140210. Bioessays. 1992. PMID: 1575712 Review.
Cited by
-
Update on the Protein Homeostasis Network in Bacillus subtilis.Front Microbiol. 2022 Mar 8;13:865141. doi: 10.3389/fmicb.2022.865141. eCollection 2022. Front Microbiol. 2022. PMID: 35350626 Free PMC article. Review.
-
CwlQ Is Required for Swarming Motility but Not Flagellar Assembly in Bacillus subtilis.J Bacteriol. 2021 Apr 21;203(10):e00029-21. doi: 10.1128/JB.00029-21. Print 2021 Apr 21. J Bacteriol. 2021. PMID: 33649146 Free PMC article.
-
McsA and B mediate the delocalization of competence proteins from the cell poles of Bacillus subtilis.Mol Microbiol. 2009 Apr;72(1):202-15. doi: 10.1111/j.1365-2958.2009.06636.x. Epub 2009 Feb 17. Mol Microbiol. 2009. PMID: 19226326 Free PMC article.
-
Roles for ClpXP in regulating the circadian clock in Synechococcus elongatus.Proc Natl Acad Sci U S A. 2018 Aug 14;115(33):E7805-E7813. doi: 10.1073/pnas.1800828115. Epub 2018 Jul 30. Proc Natl Acad Sci U S A. 2018. PMID: 30061418 Free PMC article.
-
Spatiotemporally regulated proteolysis to dissect the role of vegetative proteins during Bacillus subtilis sporulation: cell-specific requirement of σH and σA.Mol Microbiol. 2018 Apr;108(1):45-62. doi: 10.1111/mmi.13916. Epub 2018 Feb 12. Mol Microbiol. 2018. PMID: 29363854 Free PMC article.
References
-
- Ben-Yehuda, S., M. Fujita, X. S. Liu, B. Gorbatyuk, D. Skoko, J. Yan, J. F. Marko, J. S. Liu, P. Eichenberger, D. Z. Rudner, and R. Losick. 2005. Defining a centromere-like element in Bacillus subtilis by identifying the binding sites for the chromosome-anchoring protein RacA. Mol. Cell 17773-782. - PubMed
-
- Derre, I., G. Rapoport, K. Devine, M. Rose, and T. Msadek. 1999. ClpE, a novel type of HSP100 ATPase, is part of the CtsR heat shock regulon of Bacillus subtilis. Mol. Microbiol. 32581-593. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
