

Modulation of actin structure and function by phosphorylation of Tyr-53 and profilin binding
- PMID: 18689676
- PMCID: PMC2575267
- DOI: 10.1073/pnas.0805852105
Modulation of actin structure and function by phosphorylation of Tyr-53 and profilin binding
Abstract
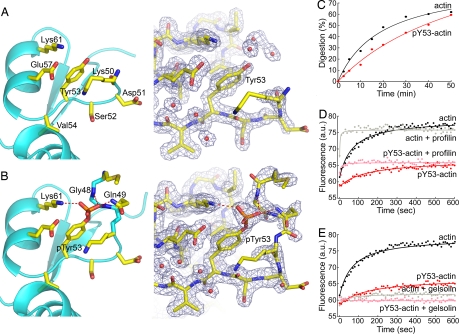
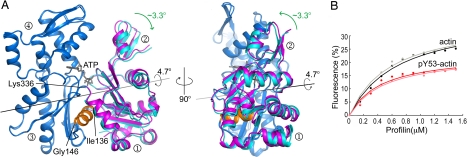
On starvation, Dictyostelium cells aggregate to form multicellular fruiting bodies containing spores that germinate when transferred to nutrient-rich medium. This developmental cycle correlates with the extent of actin phosphorylation at Tyr-53 (pY53-actin), which is low in vegetative cells but high in viable mature spores. Here we describe high-resolution crystal structures of pY53-actin and unphosphorylated actin in complexes with gelsolin segment 1 and profilin. In the structure of pY53-actin, the phosphate group on Tyr-53 makes hydrogen-bonding interactions with residues of the DNase I-binding loop (D-loop) of actin, resulting in a more stable conformation of the D-loop than in the unphosphorylated structures. A more rigidly folded D-loop may explain some of the previously described properties of pY53-actin, including its increased critical concentration for polymerization, reduced rates of nucleation and pointed end elongation, and weak affinity for DNase I. We show here that phosphorylation of Tyr-53 inhibits subtilisin cleavage of the D-loop and reduces the rate of nucleotide exchange on actin. The structure of profilin-Dictyostelium-actin is strikingly similar to previously determined structures of profilin-beta-actin and profilin-alpha-actin. By comparing this representative set of profilin-actin structures with other structures of actin, we highlight the effects of profilin on the actin conformation. In the profilin-actin complexes, subdomains 1 and 3 of actin close around profilin, producing a 4.7 degrees rotation of the two major domains of actin relative to each other. As a result, the nucleotide cleft becomes moderately more open in the profilin-actin complex, probably explaining the stimulation of nucleotide exchange on actin by profilin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Pollard TD, Borisy GG. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–465. - PubMed
-
- Engqvist-Goldstein AE, Drubin DG. Actin assembly and endocytosis: From yeast to mammals. Annu Rev Cell Dev Biol. 2003;19:287–332. - PubMed
-
- van Delft S, Verkleij AJ, Boonstra J, van Bergen en Henegouwen PM. Epidermal growth factor induces serine phosphorylation of actin. FEBS Lett. 1995;357:251–254. - PubMed
-
- Rush J, et al. Immunoaffinity profiling of tyrosine phosphorylation in cancer cells. Nat Biotechnol. 2005;23:94–101. - PubMed
-
- Kameyama K, et al. Tyrosine phosphorylation in plant bending. Nature. 2000;407:37. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
