

Regulation of apical dominance in Aspergillus nidulans hyphae by reactive oxygen species
- PMID: 18689883
- PMCID: PMC2516069
- DOI: 10.1534/genetics.108.089318
Regulation of apical dominance in Aspergillus nidulans hyphae by reactive oxygen species
Abstract
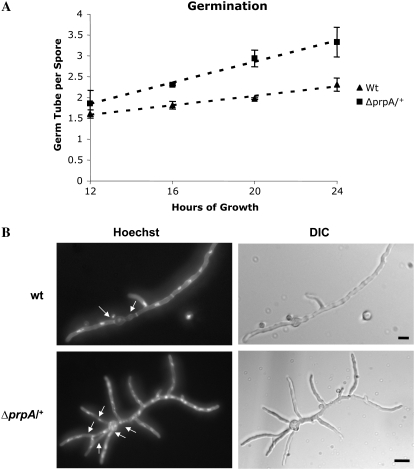
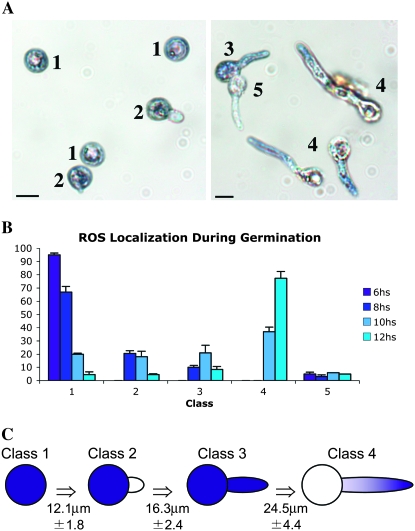
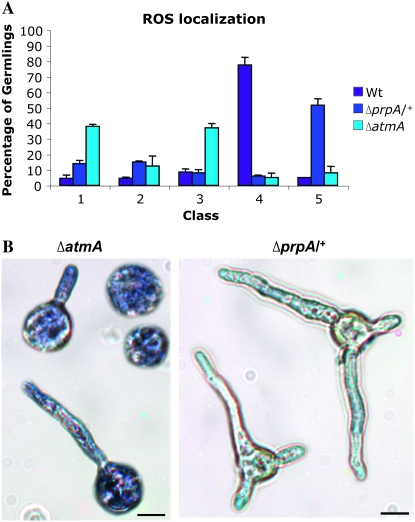
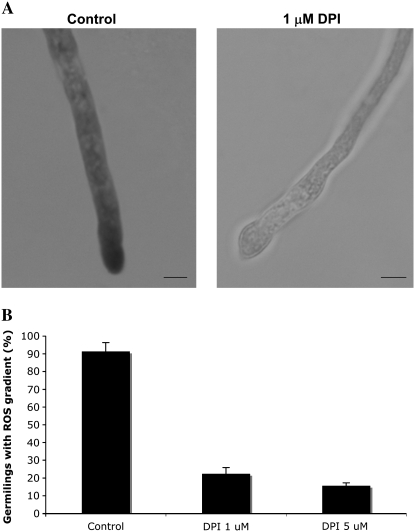
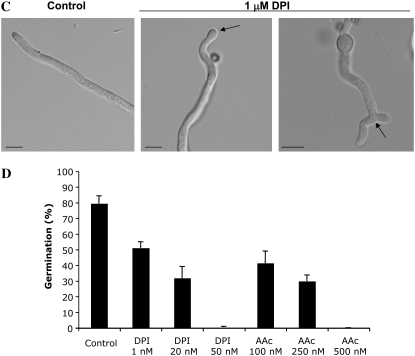
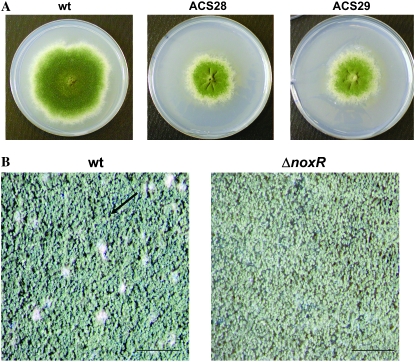
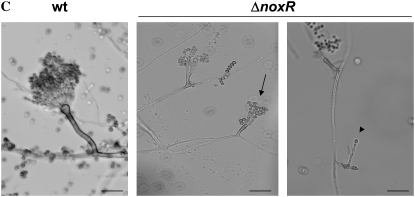
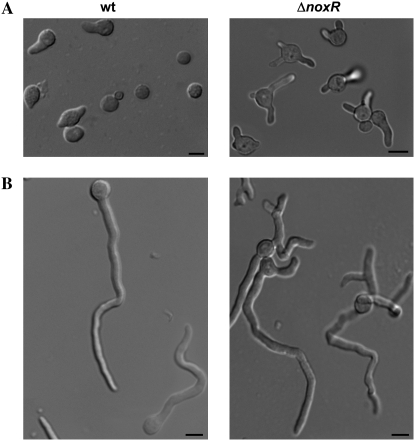
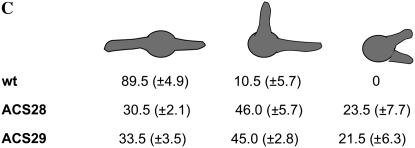
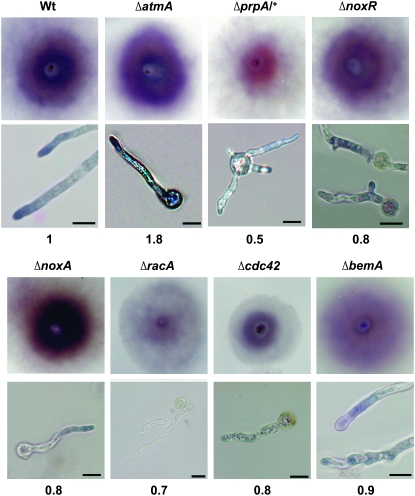
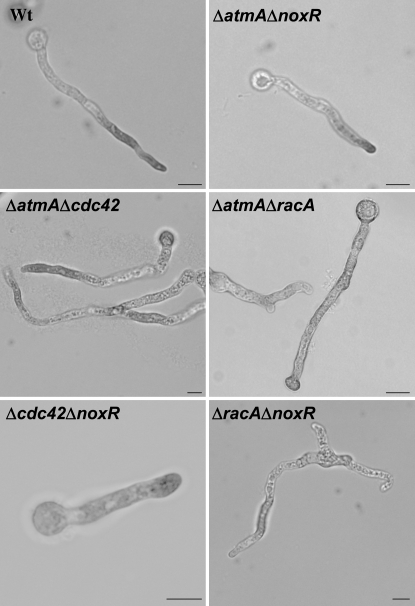
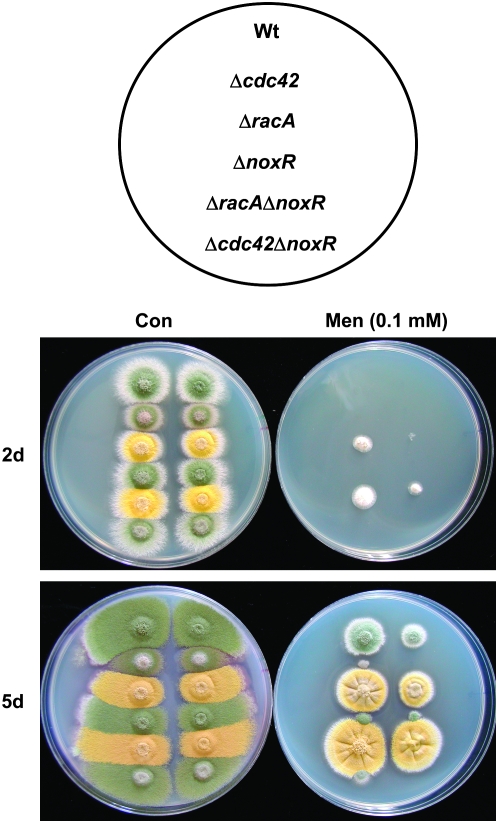
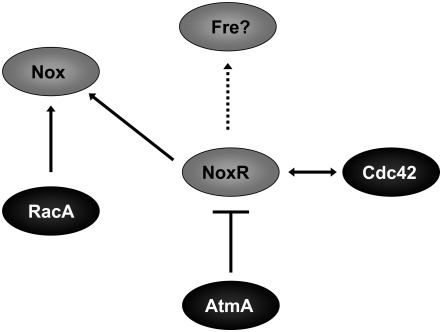
In fungal hyphae, apical dominance refers to the suppression of secondary polarity axes in the general vicinity of a growing hyphal tip. The mechanisms underlying apical dominance remain largely undefined, although calcium signaling may play a role. Here, we describe the localized accumulation of reactive oxygen species (ROS) in the apical region of Aspergillus nidulans hyphae. Our analysis of atmA (ATM) and prpA (PARP) mutants reveals a correlation between localized production of ROS and enforcement of apical dominance. We also provide evidence that NADPH oxidase (Nox) or related flavoproteins are responsible for the generation of ROS at hyphal tips and characterize the roles of the potential Nox regulators NoxR, Rac1, and Cdc42 in this process. Notably, our genetic analyses suggest that Rac1 activates Nox, whereas NoxR and Cdc42 may function together in a parallel pathway that regulates Nox localization. Moreover, the latter pathway may also include Bem1, which we propose represents a p40phox analog in fungi. Collectively, our results support a model whereby localized Nox activity generates a pool of ROS that defines a dominant polarity axis at hyphal tips.
Figures
References
-
- Abdallah, Y., D. Gligorievski, S. A. Kasseckert, L. Dieterich, M. Schafer et al., 2007. The role of poly(ADP-ribose) polymerase (PARP) in the autonomous proliferative response of endothelial cells to hypoxia. Cardiovasc. Res. 73 568–574. - PubMed
-
- Aguirre, J., M. Rios-Momberg, D. Hewitt and W. Hansberg, 2005. Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 13 111–118. - PubMed
-
- Arimura, N., and K. Kaibuchi, 2005. Key regulators in neuronal polarity. Neuron 48 881–884. - PubMed
-
- Barzilai, A., and K. Yamamoto, 2004. DNA damage responses to oxidative stress. DNA Repair 3 1109–1115. - PubMed
-
- Barzilai, A., G. Rotman and Y. Shiloh, 2002. ATM deficiency and oxidative stress: a new dimension of defective response to DNA damage. DNA Repair 1 3–25. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
