

Global analysis of the meiotic crossover landscape
- PMID: 18691940
- PMCID: PMC2628562
- DOI: 10.1016/j.devcel.2008.07.006
Global analysis of the meiotic crossover landscape
Abstract
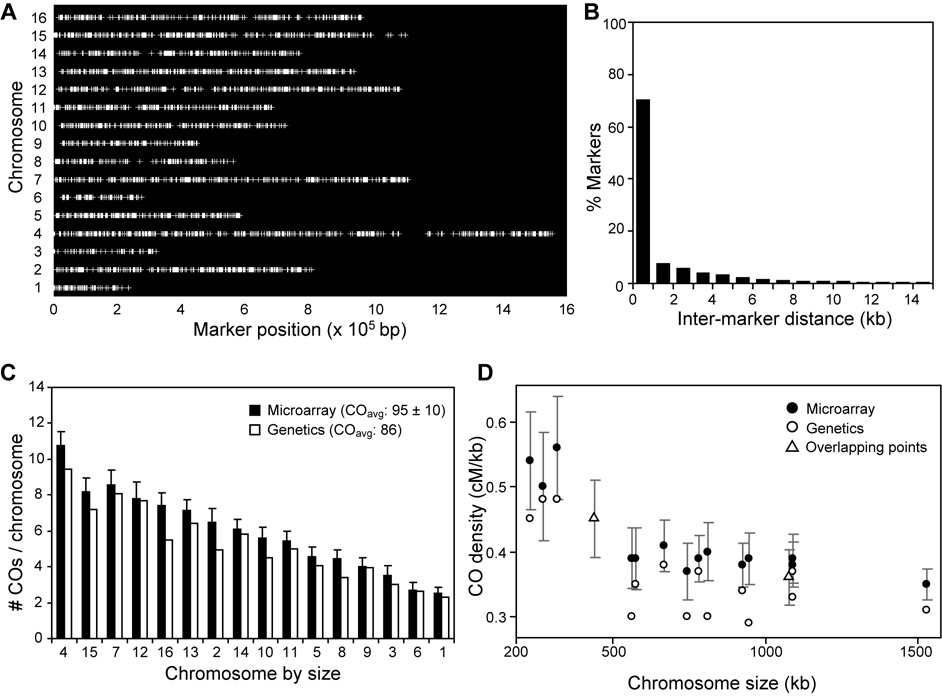
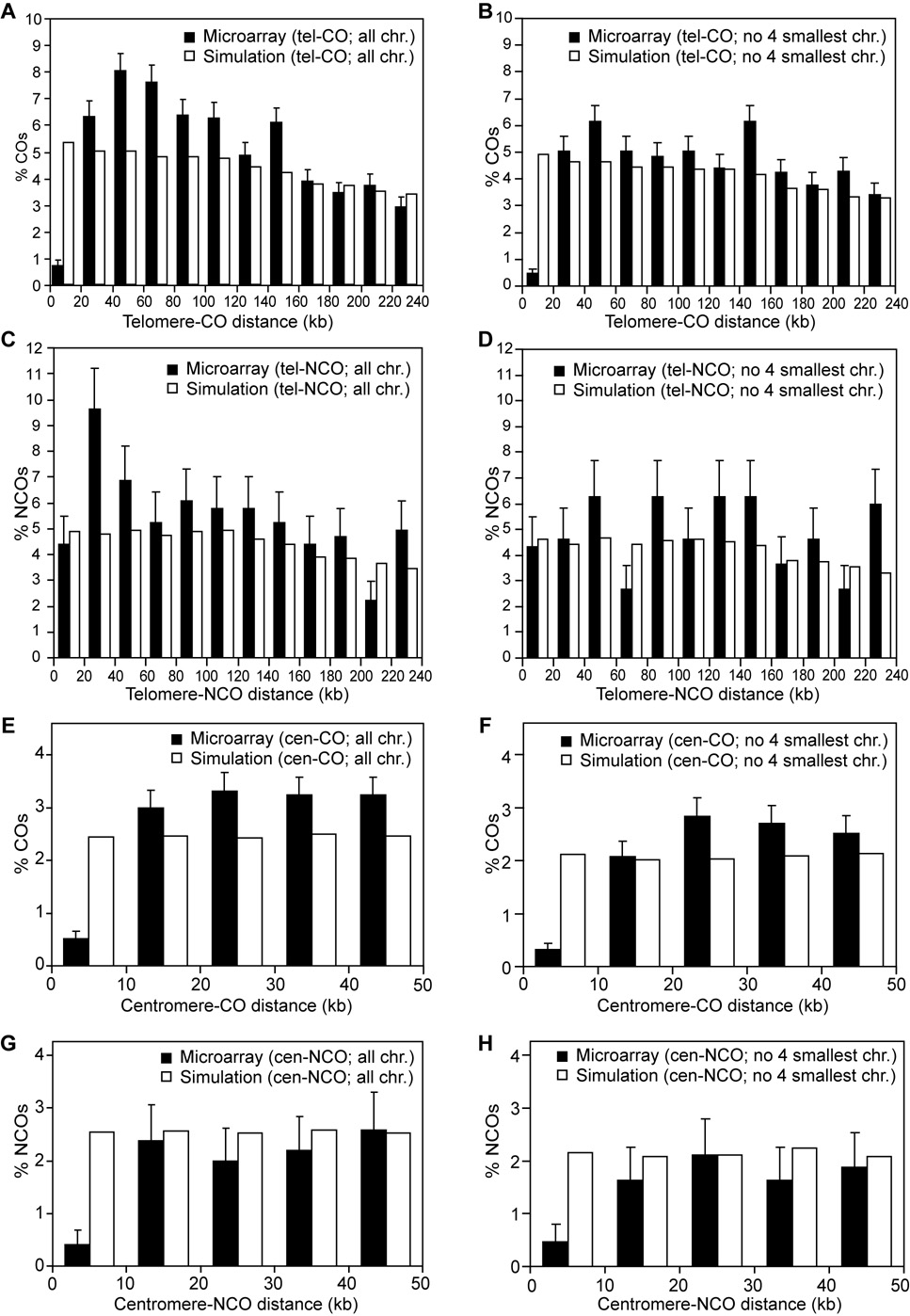
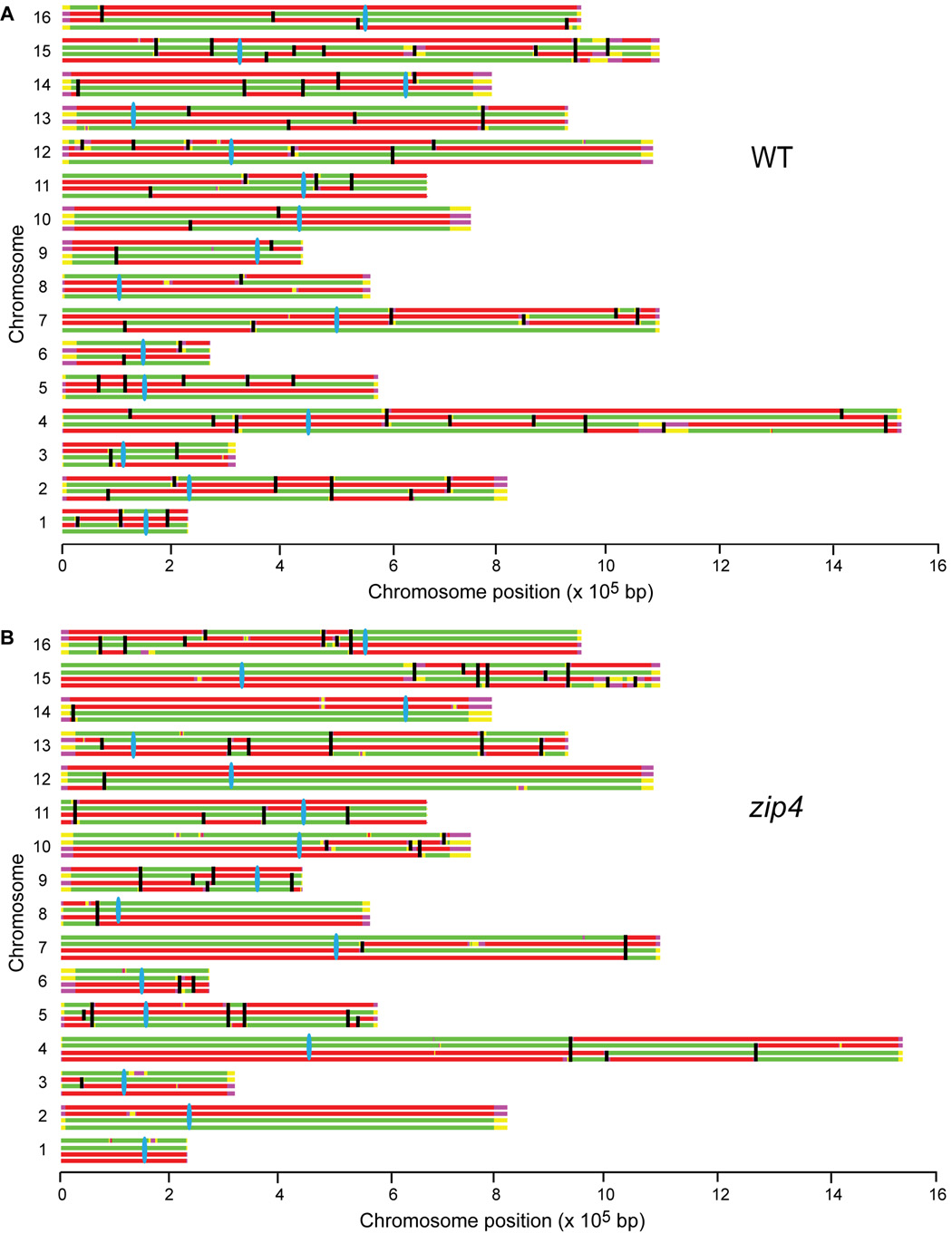
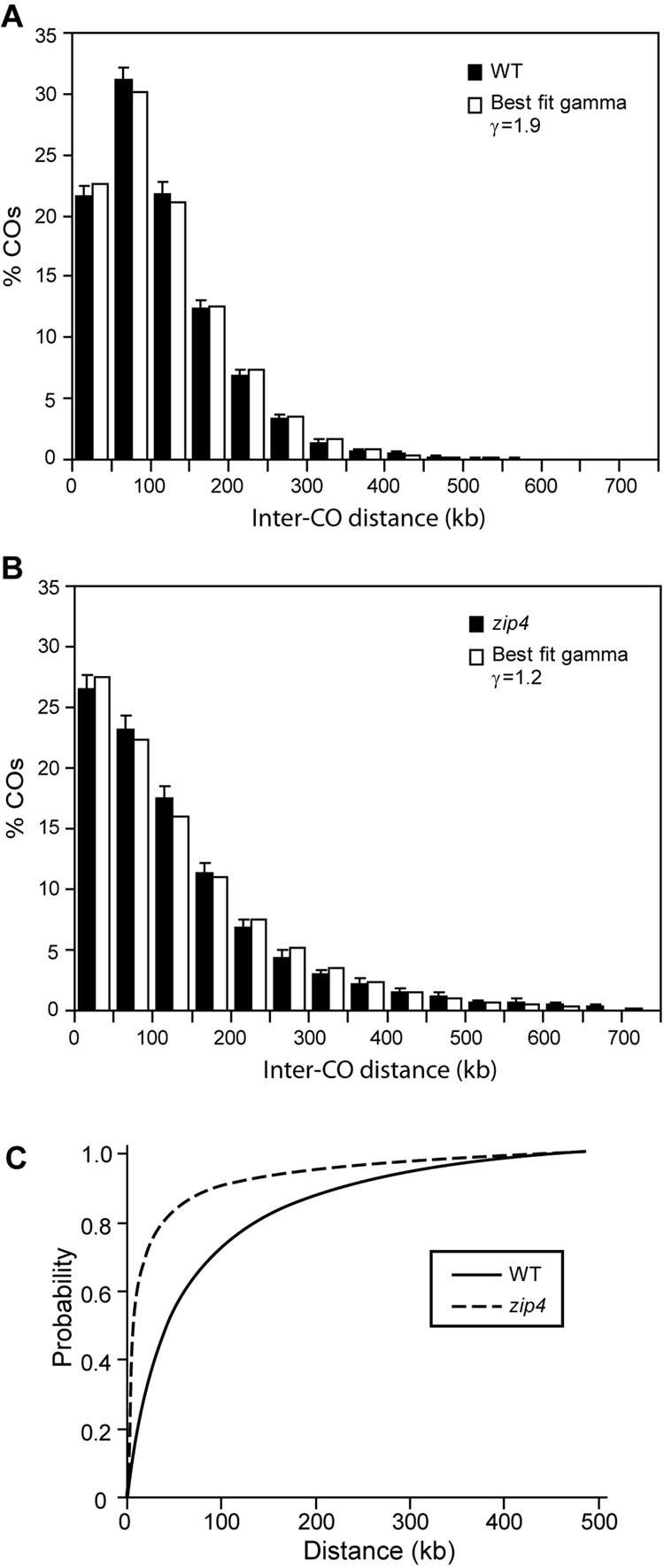
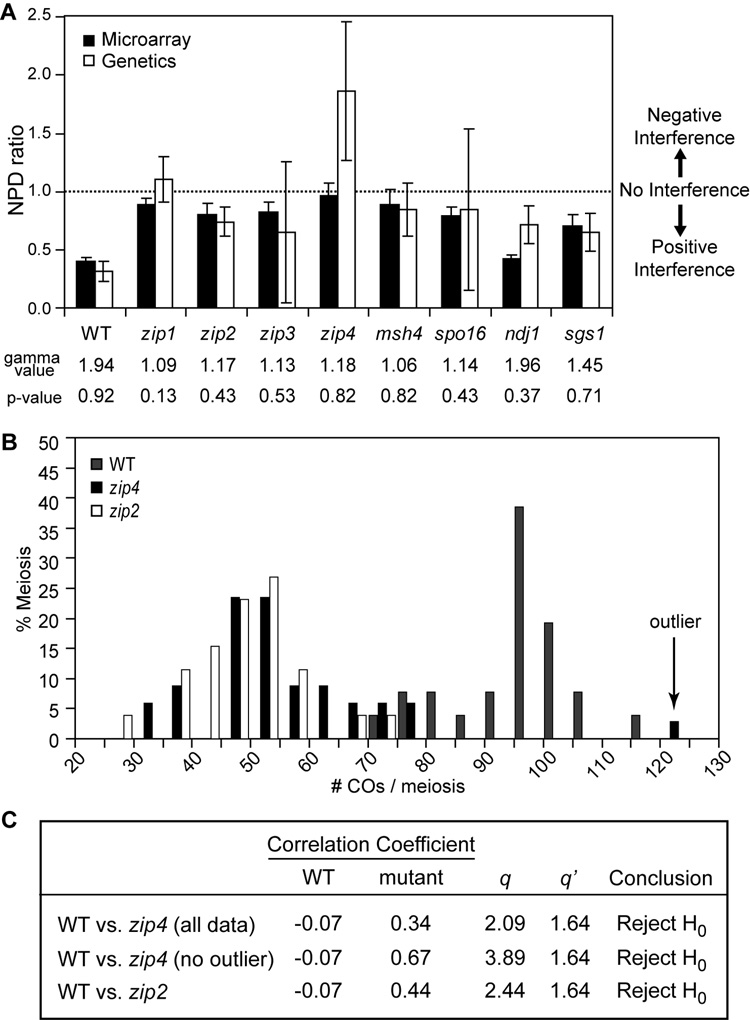
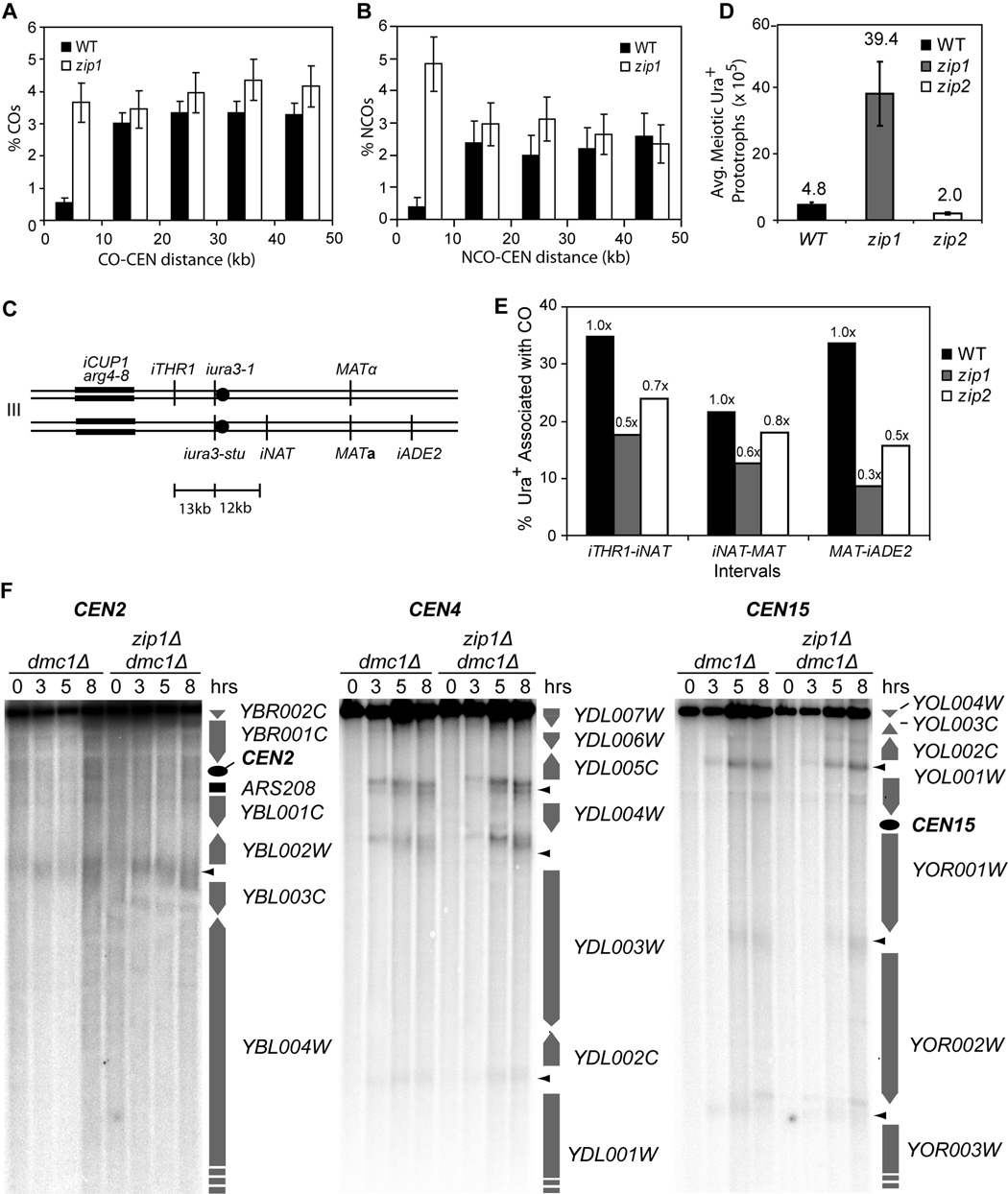
Tight control of the number and distribution of crossovers is of great importance for meiosis. Crossovers establish chiasmata, which are physical connections between homologous chromosomes that provide the tension necessary to align chromosomes on the meiotic spindle. Understanding the mechanisms underlying crossover control has been hampered by the difficulty in determining crossover distributions. Here, we present a microarray-based method to analyze multiple aspects of crossover control simultaneously and rapidly, at high resolution, genome-wide, and on a cell-by-cell basis. Using this approach, we show that loss of interference in zip2 and zip4/spo22 mutants is accompanied by a reduction in crossover homeostasis, thus connecting these two levels of crossover control. We also provide evidence to suggest that repression of crossing over at telomeres and centromeres arises from different mechanisms. Lastly, we uncover a surprising role for the synaptonemal complex component Zip1 in repressing crossing over at the centromere.
Figures
Comment in
-
Probing meiotic recombination decisions.Dev Cell. 2008 Sep;15(3):331-332. doi: 10.1016/j.devcel.2008.08.009. Dev Cell. 2008. PMID: 18804427
References
-
- Agarwal S, Roeder GS. Zip3 provides a link between recombination enzymes and synaptonemal complex proteins. Cell. 2000;102:245–255. - PubMed
-
- Baudat F, de Massy B. Regulating double-stranded DNA break repair towards crossover or non-crossover during mammalian meiosis. Chromosome Res. 2007;15:565–577. - PubMed
-
- Bishop DK, Park D, Xu L, Kleckner N. DMC1: a meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex formation, and cell cycle progression. Cell. 1992;69:439–456. - PubMed
-
- Blitzblau HG, Bell GW, Rodriguez J, Bell SP, Hochwagen A. Mapping of Meiotic Single-Stranded DNA Reveals Double-Strand-Break Hotspots near Centromeres and Telomeres. Curr Biol. 2007a - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
