

A cyclin D1/microRNA 17/20 regulatory feedback loop in control of breast cancer cell proliferation
- PMID: 18695042
- PMCID: PMC2500136
- DOI: 10.1083/jcb.200801079
A cyclin D1/microRNA 17/20 regulatory feedback loop in control of breast cancer cell proliferation
Abstract
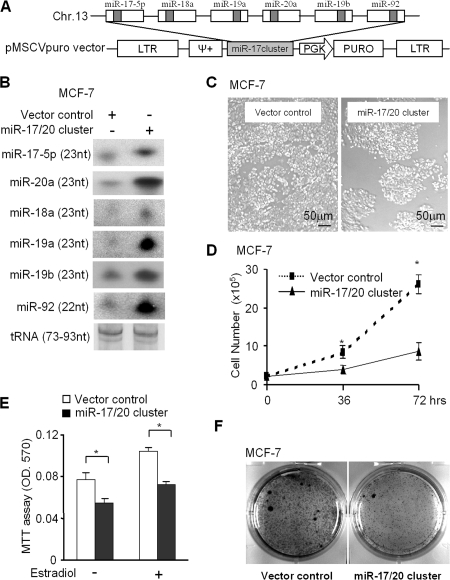
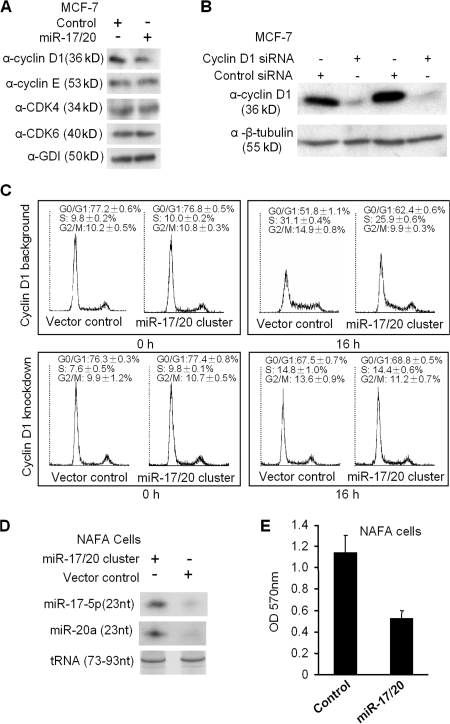
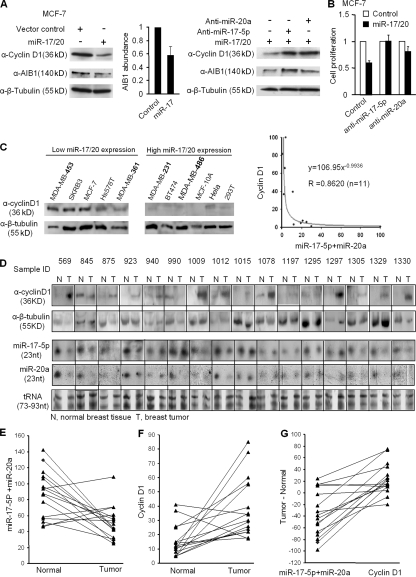
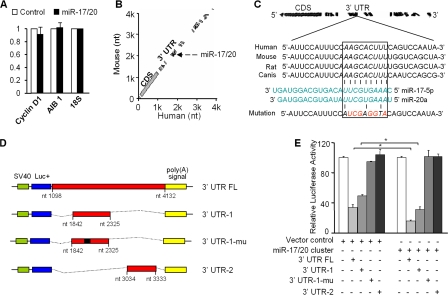
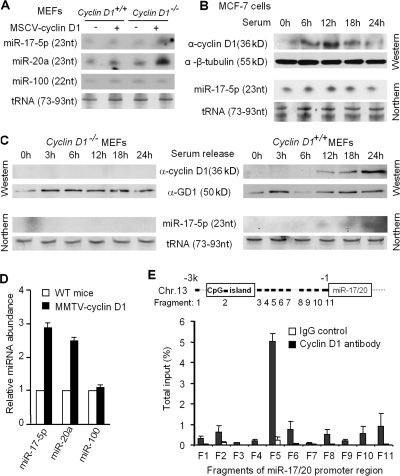
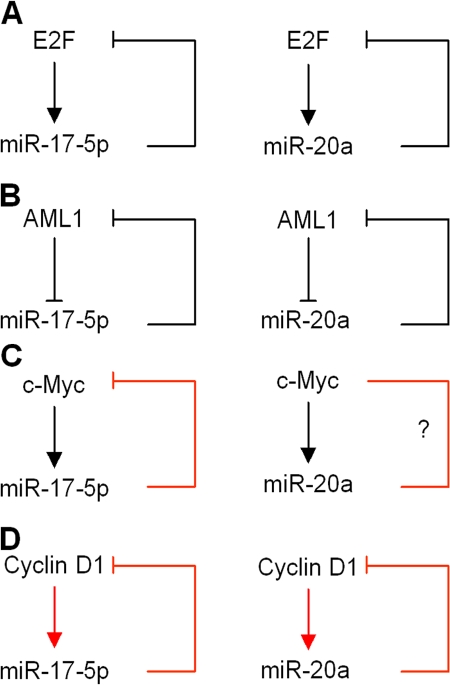
Decreased expression of specific microRNAs (miRNAs) occurs in human tumors, which suggests a function for miRNAs in tumor suppression. Herein, levels of the miR-17-5p/miR-20a miRNA cluster were inversely correlated to cyclin D1 abundance in human breast tumors and cell lines. MiR-17/20 suppressed breast cancer cell proliferation and tumor colony formation by negatively regulating cyclin D1 translation via a conserved 3' untranslated region miRNA-binding site, thereby inhibiting serum-induced S phase entry. The cell cycle effect of miR-17/20 was abrogated by cyclin D1 siRNA and in cyclin D1-deficient breast cancer cells. Mammary epithelial cell-targeted cyclin D1 expression induced miR-17-5p and miR-20a expression in vivo, and cyclin D1 bound the miR-17/20 cluster promoter regulatory region. In summary, these studies identify a novel cyclin D1/miR-17/20 regulatory feedback loop through which cyclin D1 induces miR-17-5p/miR-20a. In turn, miR-17/20 limits the proliferative function of cyclin D1, thus linking expression of a specific miRNA cluster to the regulation of oncogenesis.
Figures
References
-
- Albanese, C., J. Johnson, G. Watanabe, N. Eklund, D. Vu, A. Arnold, and R.G. Pestell. 1995. Transforming p21ras mutants and c-Ets-2 activate the cyclin D1 promoter through distinguishable regions. J. Biol. Chem. 270:23589–23597. - PubMed
-
- Ambros, V. 2004. The functions of animal microRNAs. Nature. 431:350–355. - PubMed
-
- Calin, G.A., and C.M. Croce. 2006. MicroRNA signatures in human cancers. Nat. Rev. Cancer. 6:857–866. - PubMed
-
- Calin, G.A., C. Sevignani, C.D. Dumitru, T. Hyslop, E. Noch, S. Yendamuri, M. Shimizu, S. Rattan, F. Bullrich, M. Negrini, and C.M. Croce. 2004. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA. 101:2999–3004. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
