

Embryonic chick corneal epithelium: a model system for exploring cell-matrix interactions
- PMID: 18697222
- PMCID: PMC2754064
- DOI: 10.1002/dvdy.21637
Embryonic chick corneal epithelium: a model system for exploring cell-matrix interactions
Abstract
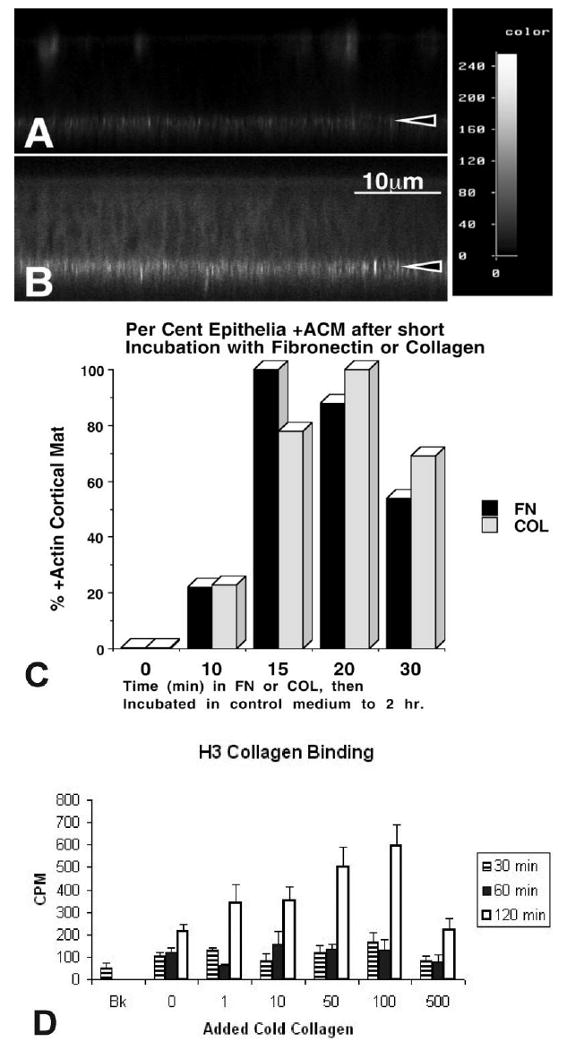
In her initial research, Elizabeth D. Hay studied amphibian limb regeneration, but later switched her focus, and for the remainder of her career addressed the role of the extracellular matrix (ECM) in regulating embryonic morphogenesis. Much of that work used the embryonic chick corneal epithelial model. This review highlights many of the discoveries that she made using this model. Hay was the first to show that embryonic corneal epithelial cells produce fibrillar collagen. Her lab was among the first to demonstrate that corneal epithelial cells respond to a collagenous substrate by increasing ECM production, and that purified ECM molecules, added to cultures of epithelial sheets, induce a reorganization of the actin cytoskeleton. These data led to the first theories of cell-matrix interactions, illustrated in a 'hands across the membrane' sketch drawn by Hay. Recent work with the epithelial sheet model system has elucidated many of the signal transduction pathways required for actin reorganization in response to the ECM. In all, this body of work has amply supported Hay's belief that the embryonic corneal epithelium is a powerful model system for exploring the role of the ECM in regulating the cytoskeleton, in directing cell migration, and in profoundly influencing cell growth and differentiation during development.
Copyright (c) 2008 Wiley-Liss, Inc.
Figures
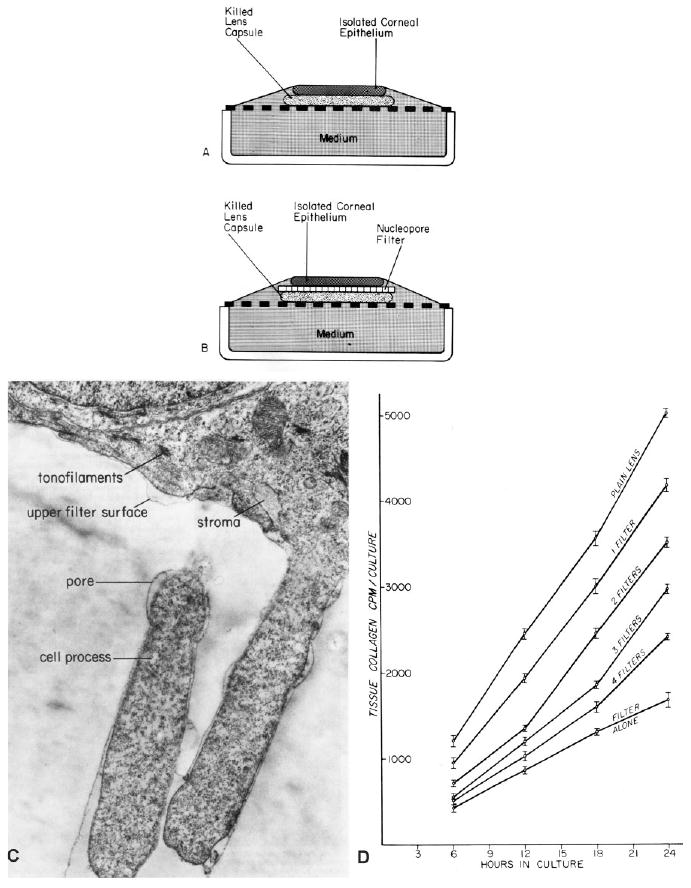
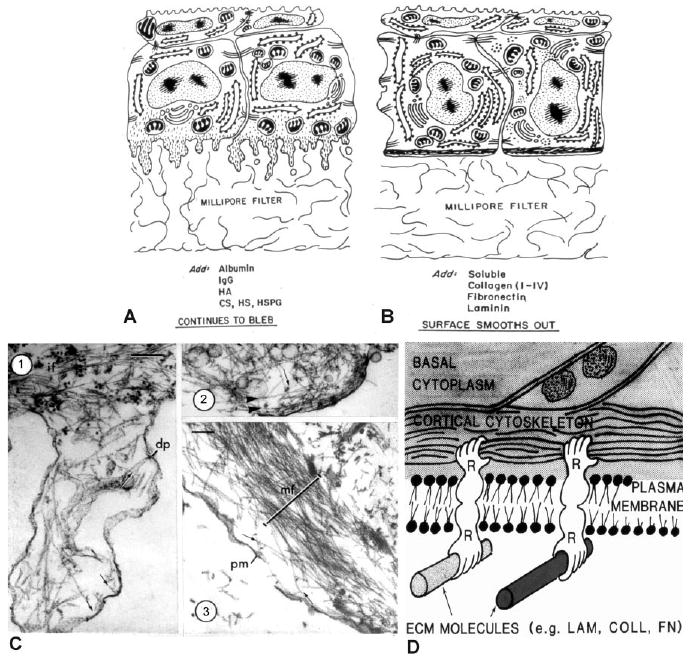
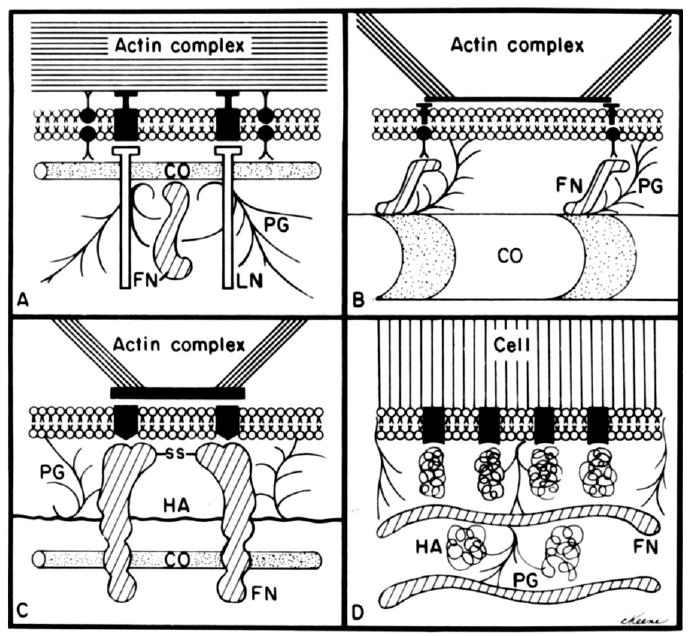
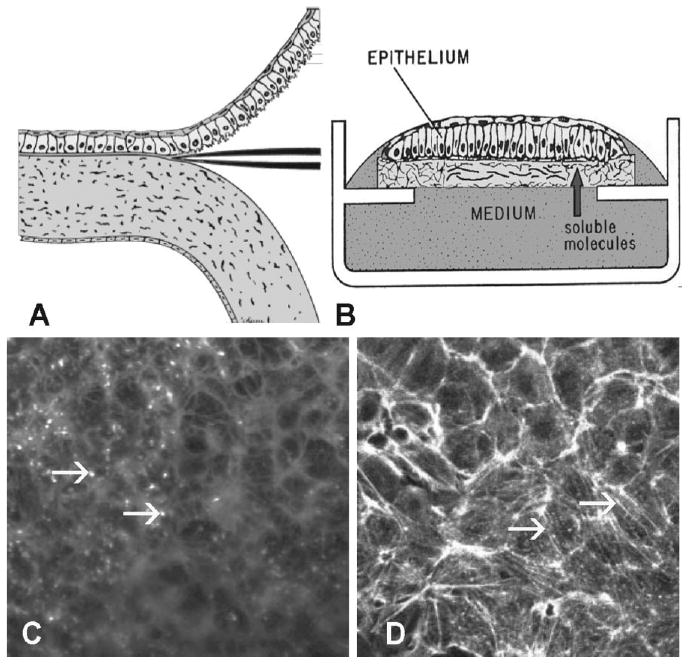
References
-
- Ali IU, Hynes RO, Ali IU, Hynes RO. Effects of LETS glycoprotein on cell motility. Cell. 1978;14:439–446. - PubMed
-
- Bernfield MR. Collagen synthesis during epitheliomesenchymal interactions. Dev Biol. 1970;22:213–231. - PubMed
-
- Coulombre AJ, Coulmbre JL. Corneal development. II. Transparency changes during rapid hydration. Am J Ophthalmol. 1958;46:276–280. discussion 281. - PubMed
-
- Dodson JW. The differentiation of epidermis. I. The interrelationship of epidermis and dermis on embryonic chicken skin. J Embryol Exp Morphol. 1967;17:83–105. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
