

From stop to start: tandem gene arrangement, copy number and trans-splicing sites in the dinoflagellate Amphidinium carterae
- PMID: 18698341
- PMCID: PMC2488372
- DOI: 10.1371/journal.pone.0002929
From stop to start: tandem gene arrangement, copy number and trans-splicing sites in the dinoflagellate Amphidinium carterae
Abstract
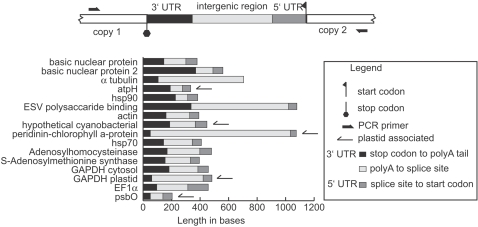
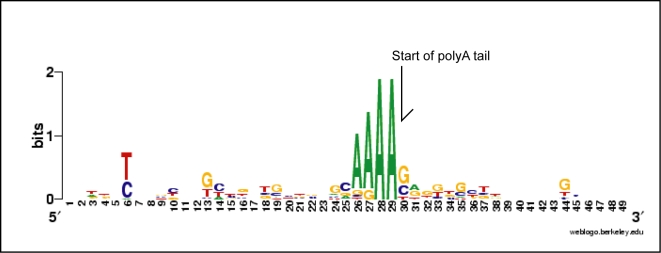
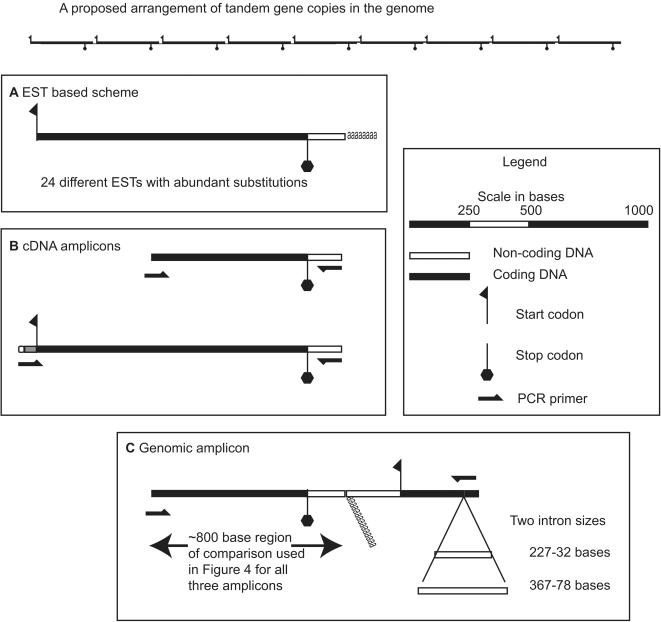
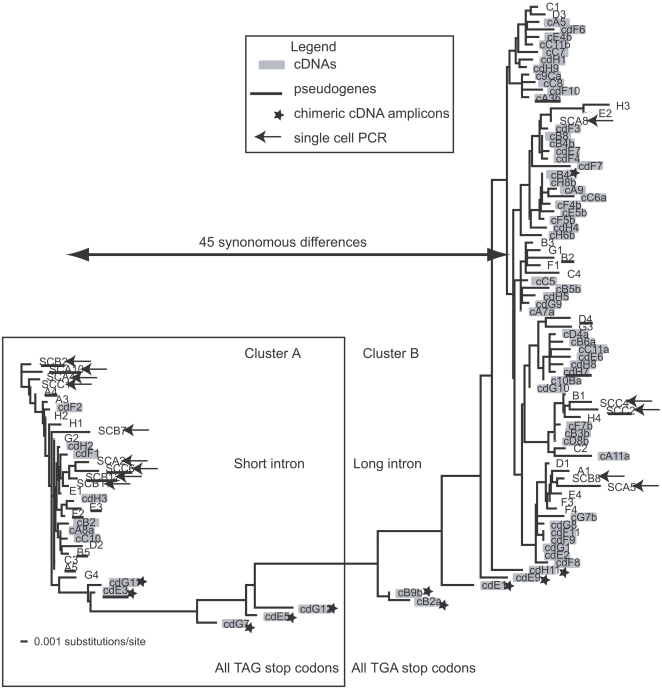
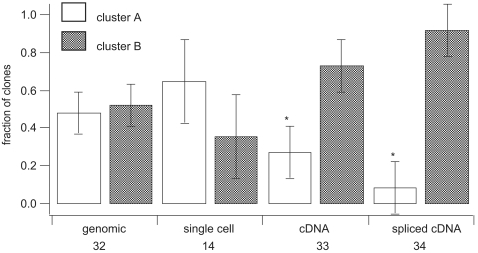
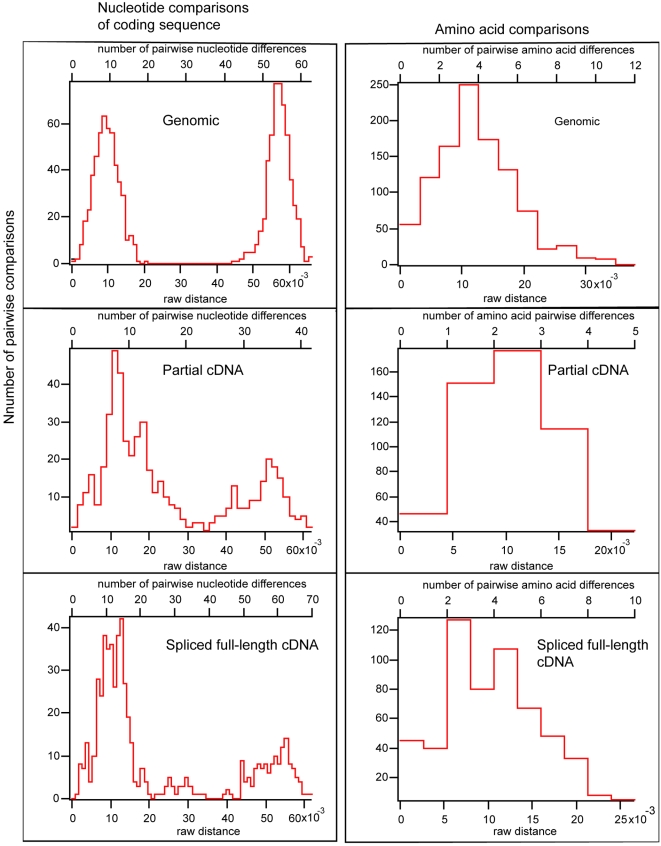
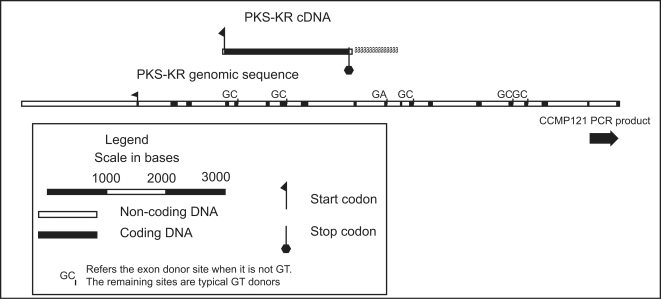
Dinoflagellate genomes present unique challenges including large size, modified DNA bases, lack of nucleosomes, and condensed chromosomes. EST sequencing has shown that many genes are found as many slightly different variants implying that many copies are present in the genome. As a preliminary survey of the genome our goal was to obtain genomic sequences for 47 genes from the dinoflagellate Amphidinium carterae. A PCR approach was used to avoid problems with large insert libraries. One primer set was oriented inward to amplify the genomic complement of the cDNA and a second primer set would amplify outward between tandem repeats of the same gene. Each gene was also tested for a spliced leader using cDNA as template. Almost all (14/15) of the highly expressed genes (i.e. those with high representation in the cDNA pool) were shown to be in tandem arrays with short intergenic spacers, and most were trans-spliced. Only two moderately expressed genes were found in tandem arrays. A polyadenylation signal was found in genomic copies containing the sequence AAAAG/C at the exact polyadenylation site and was conserved between species. Four genes were found to have a high intron density (>5 introns) while most either lacked introns, or had only one to three. Actin was selected for deeper sequencing of both genomic and cDNA copies. Two clusters of actin copies were found, separated from each other by many non-coding features such as intron size and sequence. One intron-rich gene was selected for genomic walking using inverse PCR, and was not shown to be in a tandem repeat. The first glimpse of dinoflagellate genome indicates two general categories of genes in dinoflagellates, a highly expressed tandem repeat class and an intron rich less expressed class. This combination of features appears to be unique among eukaryotes.
Conflict of interest statement
Figures

References
-
- LaJeunesse TC, Lambert G, Andersen RA, Coffroth MA, Galbraith DW. Symbiodinium (Pyrrhophyta) genome sizes (DNA content) are smallest among dinoflagellates. J Phycol. 2005;41:880–886.
-
- Rae P. Hydroxymethyluracil in eukaryote DNA: a natural feature of the pyrrophyta (dinoflagellates). Science. 1976;194:1062–1064. - PubMed
-
- Rizzo PJ, Nooden LD. Chromosomal proteins in the dinoflagellate alga Gyrodinium cohnii. Science. 1972;176:796–797. - PubMed
-
- Haapala O, Soyer -Gobillard M. Structure of dinoflagellate chromosomes. Nature. 1973;244:195–197. - PubMed
-
- Bjornland T, Tangen K. Pigmentation and morphology of a marine Gyrodinium (dinophyceae) with a major carotenoid different from peridinin and fucoxanthin. J Phycol. 1979;15:457–463.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
