

The role of bZIP transcription factors in green plant evolution: adaptive features emerging from four founder genes
- PMID: 18698409
- PMCID: PMC2492810
- DOI: 10.1371/journal.pone.0002944
The role of bZIP transcription factors in green plant evolution: adaptive features emerging from four founder genes
Abstract
Background: Transcription factors of the basic leucine zipper (bZIP) family control important processes in all eukaryotes. In plants, bZIPs are regulators of many central developmental and physiological processes including photomorphogenesis, leaf and seed formation, energy homeostasis, and abiotic and biotic stress responses. Here we performed a comprehensive phylogenetic analysis of bZIP genes from algae, mosses, ferns, gymnosperms and angiosperms.
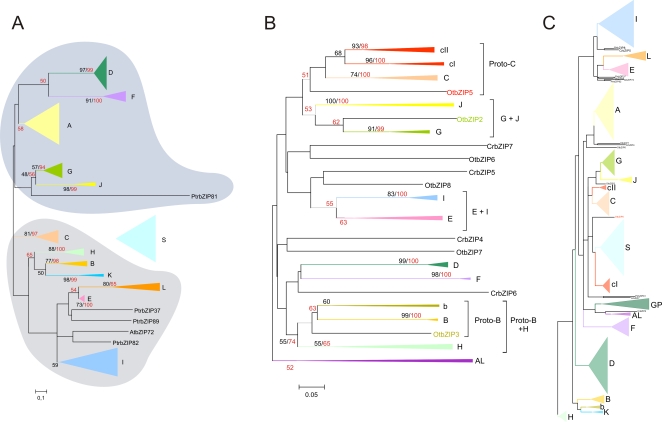
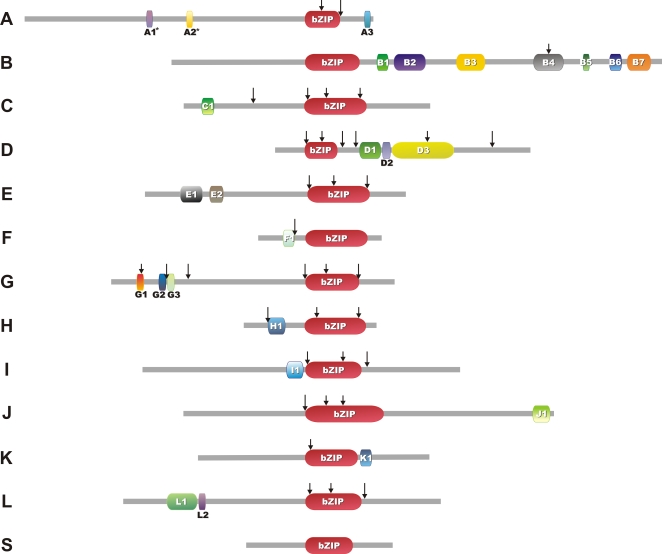
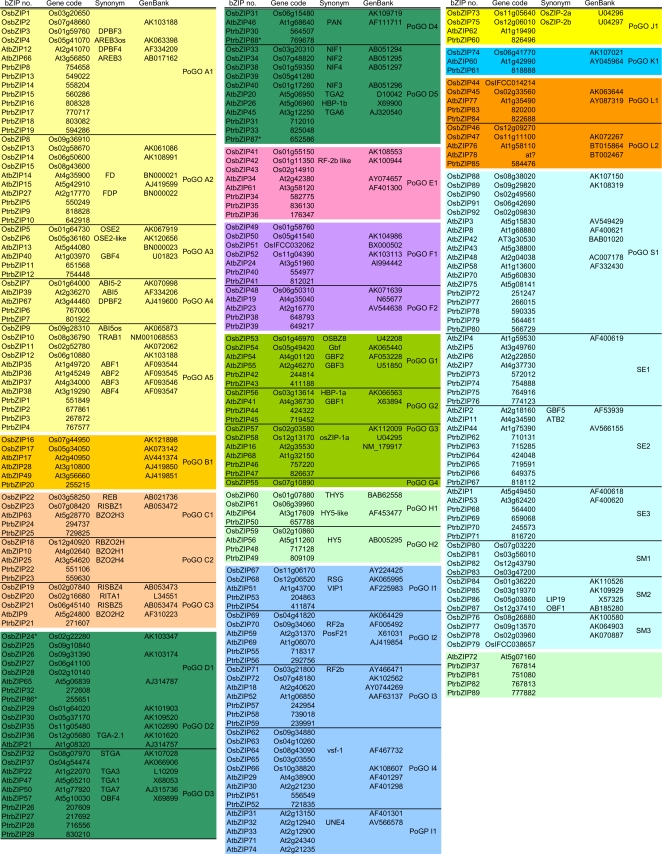
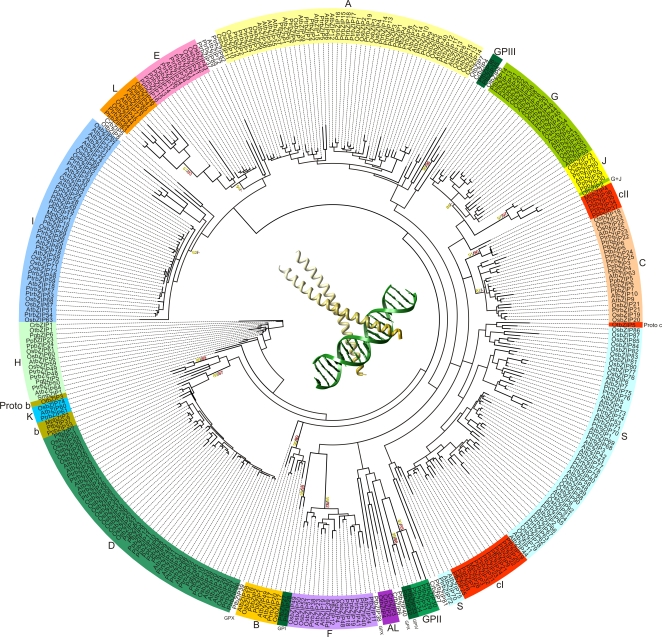
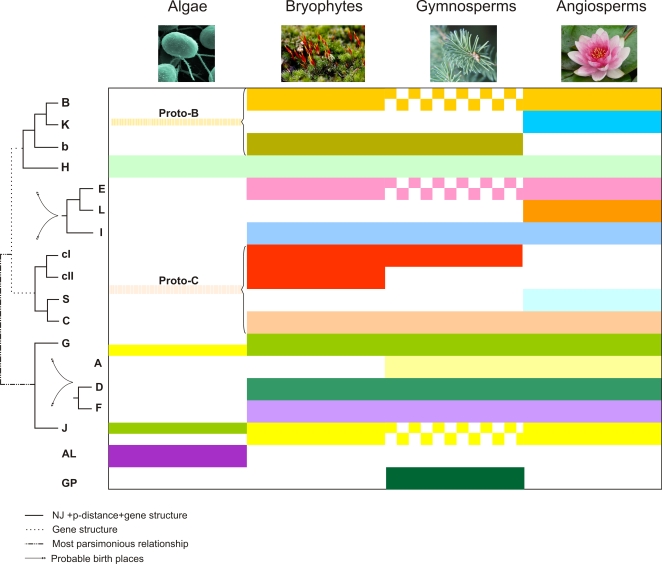
Methodology/principal findings: We identified 13 groups of bZIP homologues in angiosperms, three more than known before, that represent 34 Possible Groups of Orthologues (PoGOs). The 34 PoGOs may correspond to the complete set of ancestral angiosperm bZIP genes that participated in the diversification of flowering plants. Homologous genes dedicated to seed-related processes and ABA-mediated stress responses originated in the common ancestor of seed plants, and three groups of homologues emerged in the angiosperm lineage, of which one group plays a role in optimizing the use of energy.
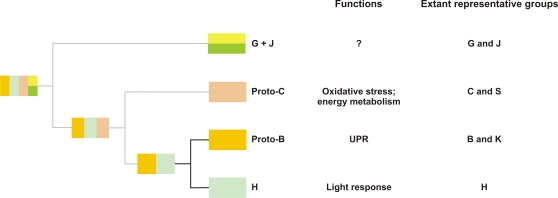
Conclusions/significance: Our data suggest that the ancestor of green plants possessed four bZIP genes functionally involved in oxidative stress and unfolded protein responses that are bZIP-mediated processes in all eukaryotes, but also in light-dependent regulations. The four founder genes amplified and diverged significantly, generating traits that benefited the colonization of new environments.
Conflict of interest statement
Figures
References
-
- Meshi T, Iwabuchi M. Plant transcription factors. Plant Cell Physiol. 1995;36:1405–1420. - PubMed
-
- Beckett D. Regulated assembly of transcription factors and control of transcription initiation. J Mol Biol. 2001;314:335–352. - PubMed
-
- Wray GA, Hahn MW, Abouheif E, Balhoff JP, Pizer M, et al. The evolution of transcriptional regulation in eukaryotes. Mol Biol Evol. 2003;20:1377–1419. - PubMed
-
- Warren AJ. Eukaryotic transcription factors. Curr Opin Struct Biol. 2002;12:107–114. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
