

Structural and functional dissection of Mif2p, a conserved DNA-binding kinetochore protein
- PMID: 18701705
- PMCID: PMC2555937
- DOI: 10.1091/mbc.e08-03-0297
Structural and functional dissection of Mif2p, a conserved DNA-binding kinetochore protein
Abstract
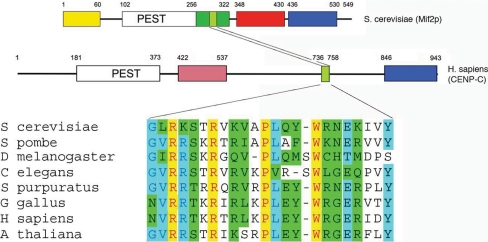
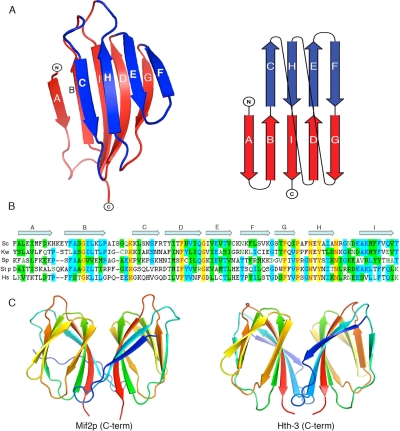
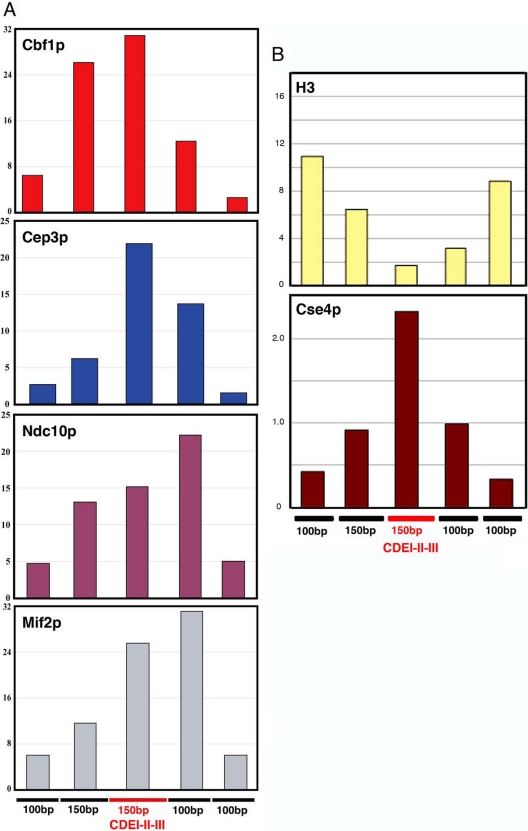
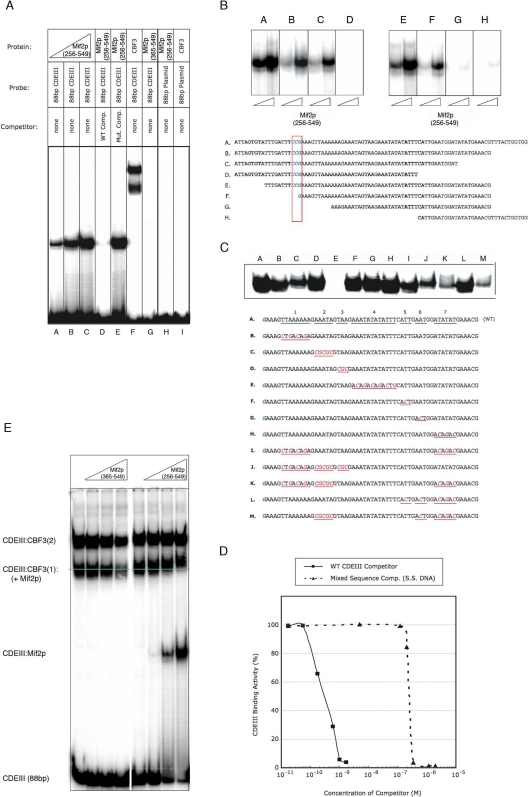
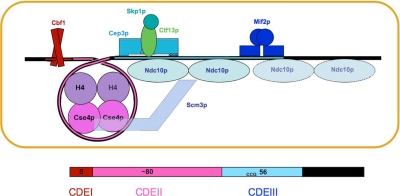
Mif2p is the budding-yeast orthologue of the mammalian centromere-binding protein CENP-C. We have mapped domains of Saccharomyces cerevisiae Mif2p and studied the phenotyptic consequences of their deletion. Using chromatin immunoprecipitation (ChIP) and electrophoretic mobility shift assays, we have further shown that Mif2p binds in the CDEIII region of the budding-yeast centromere, probably in close spatial association with Ndc10p. Moreover, ChIP experiments show that Mif2p recruits to yeast kinetochores a substantial subset of inner and outer kinetochore proteins, but not the Ndc80 or Spc105 complexes. We have determined the crystal structure of the C-terminal, dimerization domain of Mif2p. It has a "cupin" fold, extremely similar both in polypeptide chain conformation and in dimer geometry to the dimerization domain of a bacterial transcription factor. The Mif2p dimer seems to be part of an enhanceosome-like structure that nucleates kinetochore assembly in budding yeast.
Figures

Similar articles
-
The C-terminal domain of CENP-C displays multiple and critical functions for mammalian centromere formation.PLoS One. 2009 Jun 8;4(6):e5832. doi: 10.1371/journal.pone.0005832. PLoS One. 2009. PMID: 19503796 Free PMC article.
-
Crystal structure of the yeast inner kinetochore subunit Cep3p.Structure. 2007 Nov;15(11):1422-30. doi: 10.1016/j.str.2007.09.008. Structure. 2007. PMID: 17997968 Free PMC article.
-
Architecture of the budding yeast kinetochore reveals a conserved molecular core.J Cell Biol. 2003 Oct 27;163(2):215-22. doi: 10.1083/jcb.200305100. J Cell Biol. 2003. PMID: 14581449 Free PMC article.
-
Simple centromere, complex kinetochore: linking spindle microtubules and centromeric DNA in budding yeast.J Cell Biol. 2002 Apr 15;157(2):199-203. doi: 10.1083/jcb.200201052. Epub 2002 Apr 15. J Cell Biol. 2002. PMID: 11956223 Free PMC article. Review.
-
The kinetochore interaction network (KIN) of ascomycetes.Mycologia. 2016 May-Jun;108(3):485-505. doi: 10.3852/15-182. Epub 2016 Feb 23. Mycologia. 2016. PMID: 26908646 Free PMC article. Review.
Cited by
-
Where is the right path heading from the centromere to spindle microtubules?Cell Cycle. 2019 Jun;18(11):1199-1211. doi: 10.1080/15384101.2019.1617008. Epub 2019 May 20. Cell Cycle. 2019. PMID: 31075048 Free PMC article. Review.
-
Seeing is believing: our evolving view of kinetochore structure, composition, and assembly.Curr Opin Cell Biol. 2019 Oct;60:44-52. doi: 10.1016/j.ceb.2019.03.016. Epub 2019 May 9. Curr Opin Cell Biol. 2019. PMID: 31078123 Free PMC article. Review.
-
The centromere: epigenetic control of chromosome segregation during mitosis.Cold Spring Harb Perspect Biol. 2014 Nov 20;7(1):a015818. doi: 10.1101/cshperspect.a015818. Cold Spring Harb Perspect Biol. 2014. PMID: 25414369 Free PMC article. Review.
-
Cryoelectron Microscopy Structure of a Yeast Centromeric Nucleosome at 2.7 Å Resolution.Structure. 2020 Mar 3;28(3):363-370.e3. doi: 10.1016/j.str.2019.12.002. Epub 2020 Jan 30. Structure. 2020. PMID: 32004465 Free PMC article.
-
Centromere Identity and the Regulation of Chromosome Segregation.Front Cell Dev Biol. 2022 Jun 2;10:914249. doi: 10.3389/fcell.2022.914249. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35721504 Free PMC article. Review.
References
-
- Baker L. J., Dorocke J. A., Harris R. A., Timm D. E. The crystal structure of yeast thiamin pyrophosphokinase. Structure. 2001;9:539–546. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
