

The cell adhesion molecule nectin-1 is critical for normal enamel formation in mice
- PMID: 18703497
- PMCID: PMC2572697
- DOI: 10.1093/hmg/ddn243
The cell adhesion molecule nectin-1 is critical for normal enamel formation in mice
Abstract
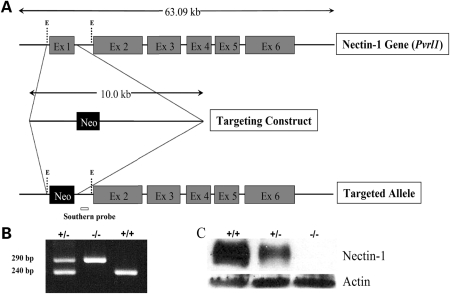
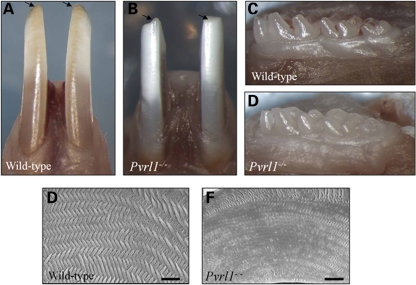
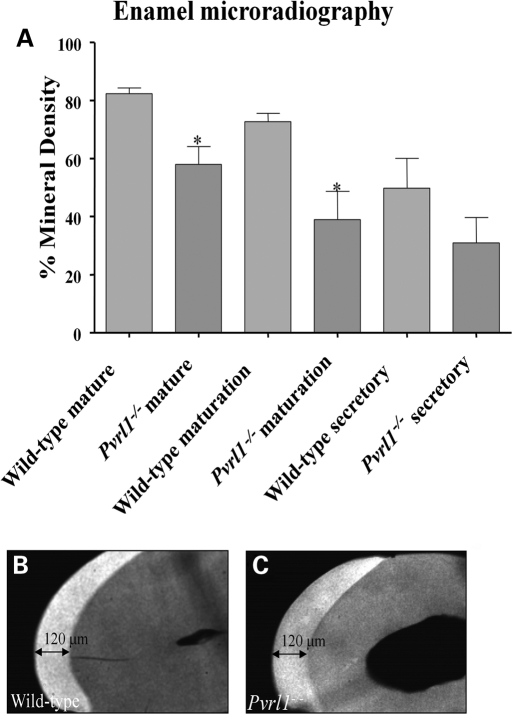
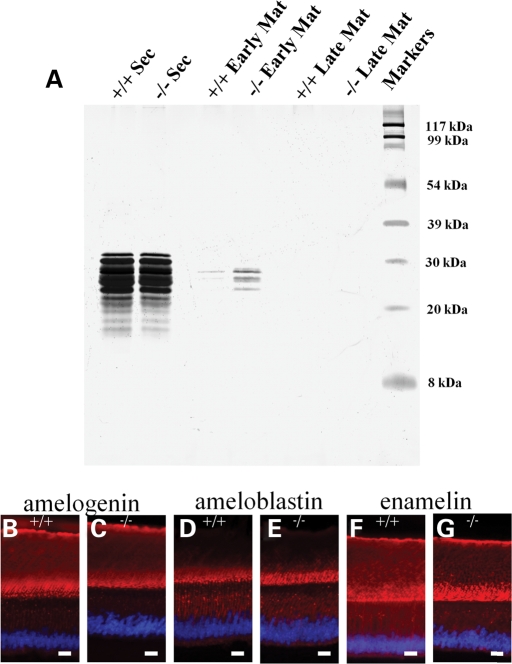
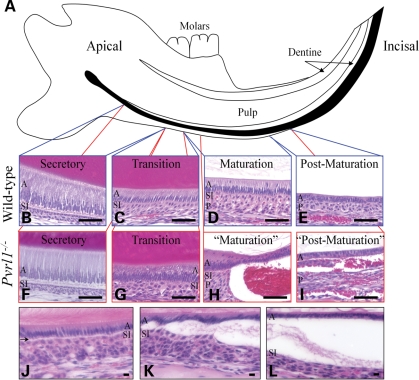
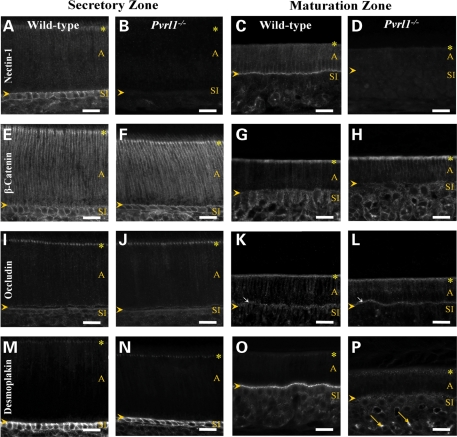
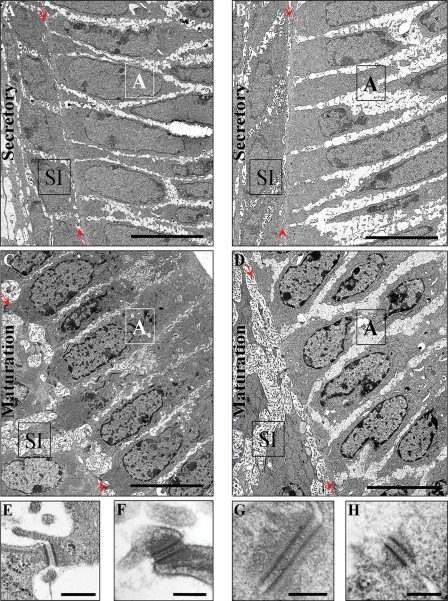
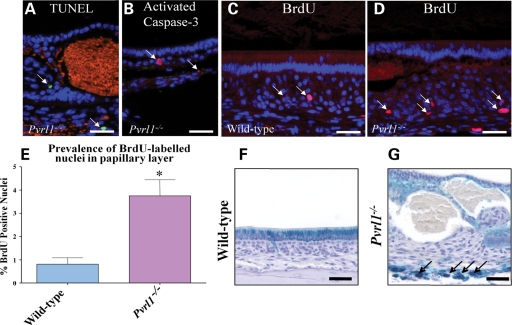
Nectin-1 is a member of a sub-family of immunoglobulin-like adhesion molecules and a component of adherens junctions. In the current study, we have shown that mice lacking nectin-1 exhibit defective enamel formation in their incisor teeth. Although the incisors of nectin-1-null mice were hypomineralized, the protein composition of the enamel matrix was unaltered. While strong immunostaining for nectin-1 was observed at the interface between the maturation-stage ameloblasts and the underlying cells of the stratum intermedium (SI), its absence in nectin-1-null mice correlated with separation of the cell layers at this interface. Numerous, large desmosomes were present at this interface in wild-type mice; however, where adhesion persisted in the mutant mice, the desmosomes were smaller and less numerous. Nectins have been shown to regulate tight junction formation; however, this is the first report showing that they may also participate in the regulation of desmosome assembly. Importantly, our results show that integrity of the SI-ameloblast interface is essential for normal enamel mineralization.
Figures
Similar articles
-
Cooperation of nectin-1 and nectin-3 is required for normal ameloblast function and crown shape development in mouse teeth.Dev Dyn. 2010 Oct;239(10):2558-69. doi: 10.1002/dvdy.22395. Dev Dyn. 2010. PMID: 21038445
-
PERP regulates enamel formation via effects on cell-cell adhesion and gene expression.J Cell Sci. 2011 Mar 1;124(Pt 5):745-54. doi: 10.1242/jcs.078071. Epub 2011 Feb 1. J Cell Sci. 2011. PMID: 21285247 Free PMC article.
-
Inhibition of Notch Signaling During Mouse Incisor Renewal Leads to Enamel Defects.J Bone Miner Res. 2016 Jan;31(1):152-62. doi: 10.1002/jbmr.2591. Epub 2015 Aug 6. J Bone Miner Res. 2016. PMID: 26179131 Free PMC article.
-
Interactions of the cell adhesion molecule nectin with transmembrane and peripheral membrane proteins for pleiotropic functions.Cell Mol Life Sci. 2008 Jan;65(2):253-63. doi: 10.1007/s00018-007-7290-9. Cell Mol Life Sci. 2008. PMID: 17928952 Free PMC article. Review.
-
The roles of nectins in cell adhesions: cooperation with other cell adhesion molecules and growth factor receptors.Curr Opin Cell Biol. 2007 Oct;19(5):593-602. doi: 10.1016/j.ceb.2007.09.007. Epub 2007 Oct 17. Curr Opin Cell Biol. 2007. PMID: 17942295 Review.
Cited by
-
Mouse models in palate development and orofacial cleft research: Understanding the crucial role and regulation of epithelial integrity in facial and palate morphogenesis.Curr Top Dev Biol. 2022;148:13-50. doi: 10.1016/bs.ctdb.2021.12.003. Epub 2022 Feb 28. Curr Top Dev Biol. 2022. PMID: 35461563 Free PMC article.
-
Hypomaturation amelogenesis imperfecta due to WDR72 mutations: a novel mutation and ultrastructural analyses of deciduous teeth.Cells Tissues Organs. 2011;194(1):60-6. doi: 10.1159/000322036. Epub 2010 Dec 29. Cells Tissues Organs. 2011. PMID: 21196691 Free PMC article.
-
p63-dependent and independent mechanisms of nectin-1 and nectin-4 regulation in the epidermis.Exp Dermatol. 2015 Feb;24(2):114-9. doi: 10.1111/exd.12593. Exp Dermatol. 2015. PMID: 25387952 Free PMC article.
-
Epithelial loss of mitochondrial oxidative phosphorylation leads to disturbed enamel and impaired dentin matrix formation in postnatal developed mouse incisor.Sci Rep. 2020 Dec 16;10(1):22037. doi: 10.1038/s41598-020-77954-7. Sci Rep. 2020. PMID: 33328493 Free PMC article.
-
Role of Nectin-1 and Herpesvirus Entry Mediator as Cellular Receptors for Herpes Simplex Virus 1 on Primary Murine Dermal Fibroblasts.J Virol. 2015 Sep;89(18):9407-16. doi: 10.1128/JVI.01415-15. Epub 2015 Jul 1. J Virol. 2015. PMID: 26136572 Free PMC article.
References
-
- Miyoshi J., Takai Y. Nectin and nectin-like molecules: biology and pathology. Am. J. Nephrol. 2007;27:590–604. - PubMed
-
- Ogita H., Takai Y. Nectins and nectin-like molecules: roles in cell adhesion, polarization, movement, and proliferation. I.U.B.M.B. Life. 2006;58:334–343. - PubMed
-
- Satoh-Horikawa K., Nakanishi H., Takahashi K., Miyahara M., Nishimura M., Tachibana K., Mizoguchi A., Takai Y. Nectin-3, a new member of immunoglobulin-like cell adhesion molecules that shows homophilic and heterophilic cell–cell adhesion activities. J. Biol. Chem. 2000;275:10291–10299. - PubMed
-
- Takai Y., Nakanishi H. Nectin and afadin: novel organizers of intercellular junctions. J. Cell Sci. 2003;116:17–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
