

Loss of cocaine locomotor response in Pitx3-deficient mice lacking a nigrostriatal pathway
- PMID: 18704092
- PMCID: PMC2752723
- DOI: 10.1038/npp.2008.117
Loss of cocaine locomotor response in Pitx3-deficient mice lacking a nigrostriatal pathway
Abstract
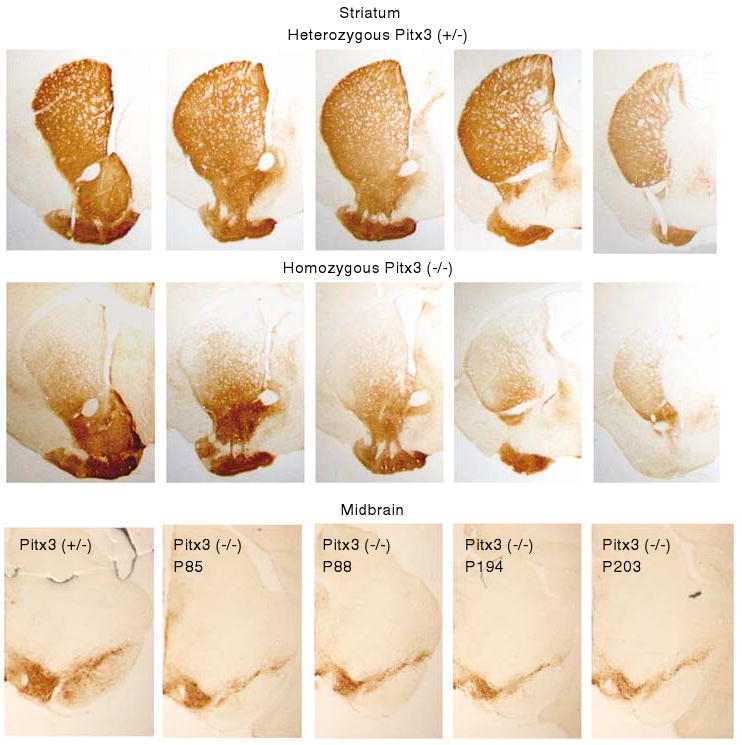
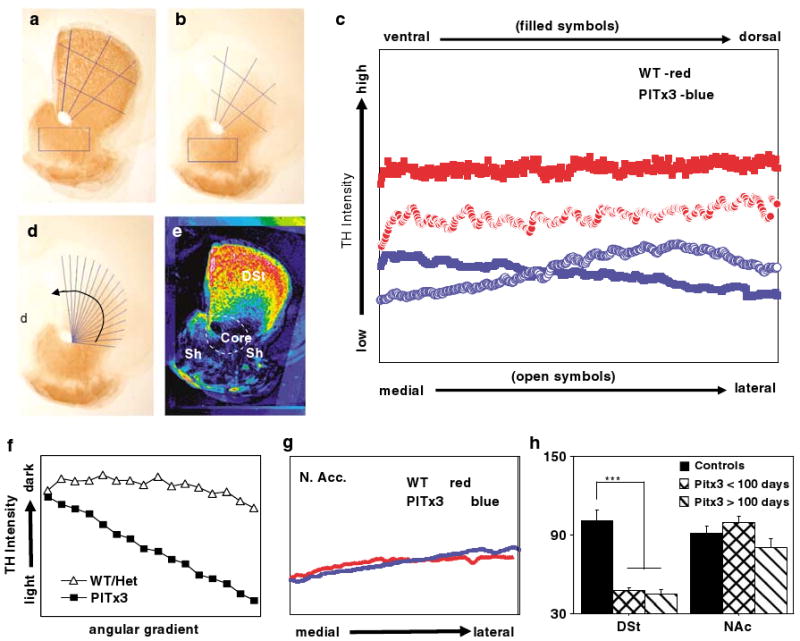
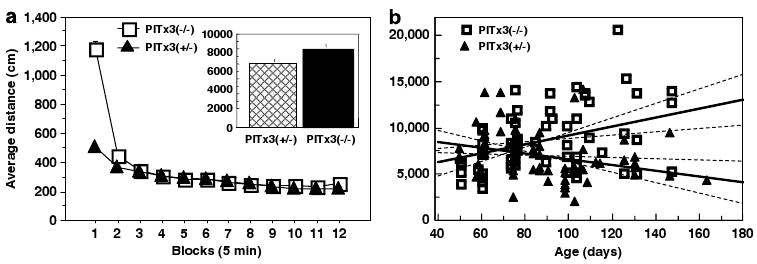
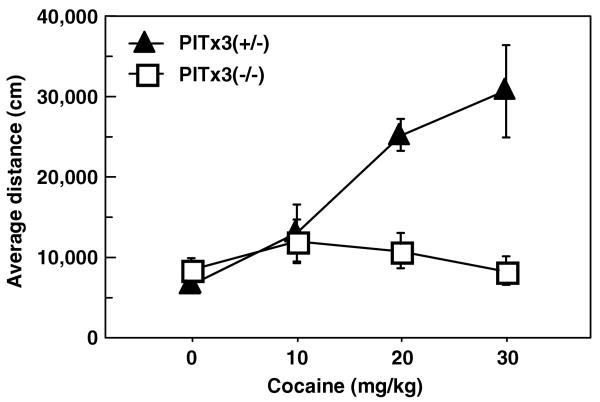
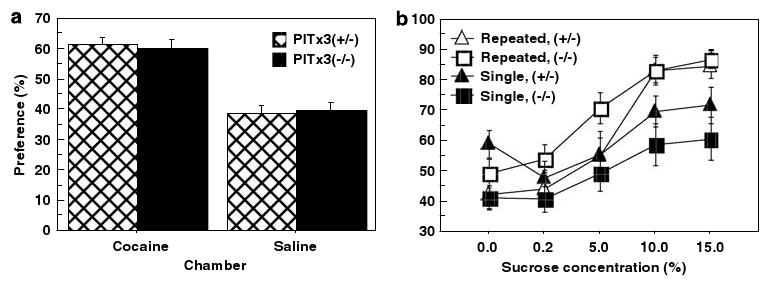
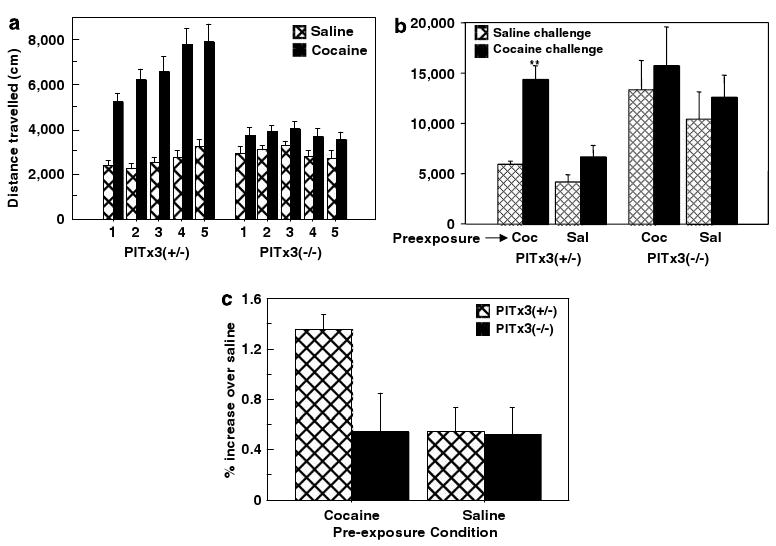
Both the dorsal and ventral striatum have been demonstrated to have a critical role in reinforcement learning and addiction. Dissecting the specific function of these striatal compartments and their associated nigrostriatal and mesoaccumbens dopamine pathways, however, has proved difficult. Previous studies using lesions to isolate the contribution of nigrostriatal and mesoaccumbens dopamine in mediating the locomotor and reinforcing effects of psychostimulant drugs have yielded inconsistent and inconclusive results. Using a naturally occurring mutant mouse line, aphakia, that lacks a nigrostriatal dopamine pathway but retains an intact mesoaccumbens pathway, we show that the locomotor activating effects of cocaine, including locomotor sensitization, are dependent on an intact nigrostriatal dopamine projection. In contrast, cocaine reinforcement, as measured by conditioned place preference and cocaine sensitization of sucrose preference, is intact in these mice. In light of the well-established role of the nucleus accumbens in mediating the effects of psychostimulants, these data suggest that the nigrostriatal pathway can act as a critical effector mechanism for the nucleus accumbens highlighting the importance of intrastriatal connectivity and providing insight into the functional architecture of the striatum.
Conflict of interest statement
Figures
References
-
- Adams JU, Careri JM, Efferen TR, Rotrosen J. Differential effects of dopamine antagonists on locomotor activity, conditioned activity and conditioned place preference induced by cocaine in rats. Behav Pharmacol. 2001;12:603–611. - PubMed
-
- Anderson SM, Pierce RC. Cocaine-induced alterations in dopamine receptor signaling: implications for reinforcement and reinstatement. Pharmacol Ther. 2005;106:389–403. - PubMed
-
- Beeler JA, Prendergast B, Zhuang X. Low amplitude entrainment of mice and the impact of circadian phase on behavior tests. Physiol Behav. 2006;87:870–880. - PubMed
-
- Belin D, Everitt BJ. Cocaine seeking habits depend upon dopamine-dependent serial connectivity linking the ventral with the dorsal striatum. Neuron. 2008;57:432–441. - PubMed
-
- Cagniard B, Beeler JA, Britt JP, McGehee DS, Marinelli M, Zhuang X. Dopamine scales performance in the absence of new learning. Neuron. 2006;51:541–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
